



Mycotic diseases are found in marine mammals worldwide, and captive marine mammals seem particularly prone to fungal infections. Cetaceans lack nasal turbinates, which may allow fungal elements to more easily enter the pulmonary system. Most infections appear to be secondary to stress, environmental compromise, or concurrent infectious disease. Some systemic mycoses have distinct geographic distributions, with many fungi ubiquitous in these environments.
Diagnosis is based on clinical signs and confirmed by identification of the organism on cytologic evaluation, biopsy, or, preferably, culture. Serologic testing may be useful in some infections. Lactophenol cotton blue wet mount preparation may render an immediate diagnosis for some of the morphologically distinct fungi. Tissue smears cleared in warm 10% potassium hydroxide can be examined to identify characteristic fruiting bodies or hyphae.
Topical medication of pinnipeds for dermatophytosis is feasible. Smaller cetaceans can be kept out of water in a sling for 2–24 hours, provided that areas of the body not being treated are kept moist; however, this technique is less than ideal due to potential discomfort for the animal. Otherwise, systemic therapy is used, including oral and intravenous routes as well as nebulization in pulmonary cases.
Aspergillosis of Marine Mammals
Fatal pulmonary aspergillosis has been diagnosed in several species of cetaceans, including bottlenose dolphins and killer whales, and in several pinnipeds, including Antarctic fur seals (Arctocephalus gazella), harbor seals, and California sea lions. In addition to disseminated aspergillosis, case reports of aspergillomas of the inner ear have been reported in free-ranging harbour porpoise, and cutaneous aspergillosis has been seen in gray seals (Halichoerus grypus) with concomitant mycobacteriosis.
The respiratory form has often been diagnosed on postmortem examination; however, increased vigilance with regular monitoring of respiratory samples from cetaceans has allowed earlier diagnosis and treatment.
Prophylactic antifungal use has also increased with anticipation of stressful situations, such as transport. Successful treatment is still difficult due to delays in treatment associated with diagnosis, invasive nature of the fungal agent, and difficulty balancing a safe and effective dosing range. Combinations of antifungal drugs such as triazoles and echinocandins may be more successful. Cutaneous lesions have responded to topical povidone-iodine with ketoconazole therapy (10 mg/kg, PO, every 24 hours).
Candidiasis of Marine Mammals

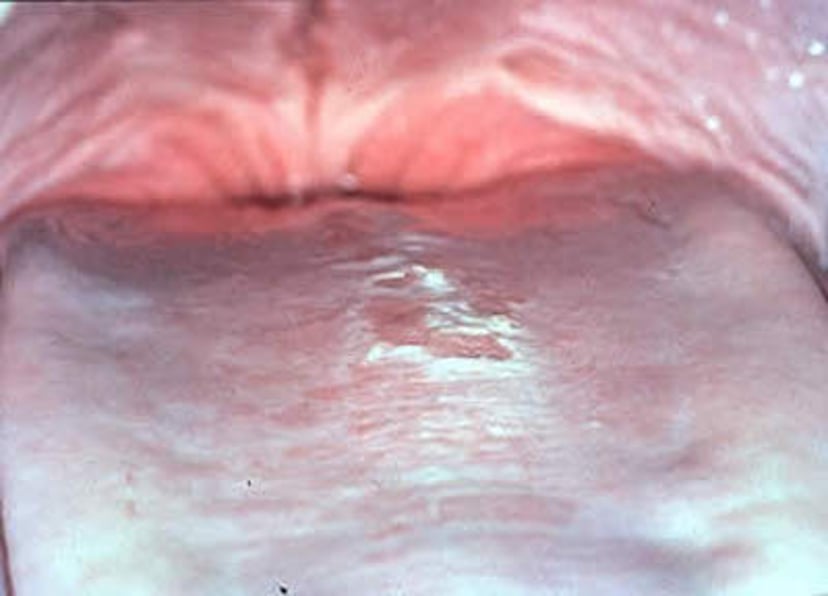
Lingual candidiasis in a bottlenose dolphin. White plaques on mucosal tissue are commonly noted with this disease.
Courtesy of Dr. James McBain.


Candidiasis in a spinner dolphin. Pale raised lesions are present around the blowhole.
Courtesy of Dr. Louise Bauck.
Candida spp are often part of the normal flora of animals, making diagnosis of actual candidiasis more challenging. This common mycotic disease in captive cetaceans occurs secondary to stress, unbalanced water disinfection with chlorines, or indiscriminate or prolonged antimicrobial therapy. Candidiasis is also reported in pinnipeds and has been found in cultures of wild cetacean blowholes and stomachs. In clinical disease in cetaceans, the lesions usually are found around body orifices. With endoscopy or at necropsy, esophageal ulcers are often found, particularly in the area of the gastroesophageal junction. In phocids, inflammation at the mucocutaneous junctions—particularly at the commissures of the mouth and around the eyes, anus, and vulva—is common, although it may also be noted in the GI tract.
Diagnosis is based on identification of the yeast on culture, cytologic evaluation, or biopsy. Early detection and treatment is usually successful. Candidiasis generally responds well to ketoconazole, itraconazole, and echinocandins; nystatin is often safe and effective for oral and GI disease. Any environmental deficits should be corrected. Supplementation with prednisolone (0.01 mg/kg, PO, every 24 hours) may be appropriate to compensate for ketoconazole inhibition of glucocorticoid production. One anecdotal report suggests a possible toxic reaction to ketoconazole in a northern elephant seal (Mirounga angustirostris).
Cryptococcosis of Marine Mammals
Another opportunistic yeast, Cryptococcus spp, has been increasingly diagnosed in the Pacific Northwest in stranded marine mammals. Organisms are usually inhaled and may be dormant before reactivating, particularly in the lungs and lymph nodes. Cryptococcus neoformans was also diagnosed in fatal advanced pulmonary disease in a bottlenose dolphin. Prolonged treatment of the dolphin with itraconazole (120 days) at routine mammalian doses was ineffective despite serum drug concentrations above the suggested therapeutic range.
Dermatophytosis of Marine Mammals
Mycotic dermatitis due to Trichophyton spp or Microsporum canis has previously responded well to topical povidone-iodine, oral griseofulvin, or both. More recently, a combination of long-term (~65 days) topical treatment with enilconazole in conjunction with oral administration of terbinafine has successfully been used to treat dermatomycosis in several sea lions.
Paracoccidioidomycosis (Lobomycosis, Lacaziosis) of Marine Mammals


Paracoccidioidomycosis (Lobomycosis), due to Paracoccidioides brasiliensisceti (formerly Loboa loboi), in an Atlantic bottlenose dolphin. Lesions are often white to gray raised, keloid-like lesions with nodules and plaques.
Courtesy of Dr. James McBain.
This disfiguring cutaneous disease is caused by infection with Paracoccidioides brasiliensisceti (formerly Lacazia loboi) in dolphins. The disease has been reported only in humans, Atlantic bottlenose dolphins, and in dolphins in Central and South America.
Culture of the organism has not yet been successful, and diagnosis is based on histopathologic findings. Differential diagnoses include sporotrichosis and chromomycosis and other fungal diseases characterized by extensive granuloma formation.
Excisional therapy and systemic antifungal drugs have been used with varying success.
Zoonotic transmission is strongly suspected in a case of a European handler of an infected dolphin. Most human cases have no history of marine mammal contact.
Systemic Mycoses of Marine Mammals
The systemic mycoses of marine mammals are a zoonotic risk, and precautions should be taken to prevent infection when handling dead and diseased animals. Cystofilobasidiales has caused fatal disease in a California sea lion. Blastomycosis has caused fatal disease in bottlenose dolphins, California sea lions, a Stellar sea lion (Eumetopias jubatus), Northern fur seals, and polar bears. Fatal systemic histoplasmosis has been reported in a captive harp seal (Pagophilus groenlandicus), a bottlenose dolphin, and a Pacific white-sided dolphin. Coccidioidomycosis has been found in bottlenose dolphins, California sea lions, and sea otters. Increasing numbers of cases are found in sea otters and sea lions.
Blastomycosis has been successfully treated with intensive management, including itraconazole (3.5 mg/kg, PO, every 24 hours for 70 days), combined with antimicrobial and supportive therapy when indicated.
Zygomycetes of Marine Mammals
Zygomycosis (or mucormycosis) is caused by numerous filamentous fungi, including Mucor, Rhizopus, Cunninghamella, Apophysomyces spp, Fusarium spp, and others and is seen increasingly often in captive marine mammals. Dermatologic conditions caused by various Fusarium spp have been reported in pygmy sperm whales (Kogia breviceps), Atlantic white-sided dolphins (Laegenorhynchus acutus), harbor seals, gray seals, California sea lions, and northern elephant seals. Mucorales have caused fatal disease in bottlenose dolphins, harbour porpoises, and harp seals. Other zygomycetes have been diagnosed as a cause of fatal disseminated disease in various species of marine mammals. These infections are often considered diseases of debilitated animals; the underlying cause of the low host resistance to these opportunistic infections must be corrected if therapy is to be successful.
Diagnosis is based on culture or organism identification from biopsy.
Cases have responded to ketoconazole (5 mg/kg, PO, every 24 hours for 10 days), fluconazole (0.5 mg/kg, PO, every 12 hours for 21 days), or itraconazole (1 mg/kg, PO, every 24 hours for 120 days). Localized fusariomycosis has been successfully treated in a captive beluga whale (Delphinapterus leucas) by use of voriconazole after surgical debridement.
Amphotericin B has been the therapy of choice for zygomycete infections, but newer imidazoles such as posaconazole may have clinical efficacy despite laboratory tests of resistance. Combined therapies may also be more effective.





