




Porcine circoviruses have been associated with multiple disease conditions in pigs, including postweaning multisystemic wasting syndrome and reproductive disorders. Virus has been detected in all body secretions. Depending on the specific type of circovirus, signs may include weight loss, late-term abortions and stillbirths, and a necrotizing vasculitis. Control of PCV-2 is by vaccination.
Porcine circovirus diseases have traditionally been defined as those associated with porcine circovirus 2 (PCV-2) infection. However, this terminology should be revised because the advent of the novel porcine circovirus 3 (PCV-3) may expand the clinicopathologic outcome caused by porcine circovirus infections. Porcine circovirus 1 (PCV-1) is still considered a nonpathogenic agent for swine.
Historically, PCV-1 was described as a noncytopathogenic, picornavirus-like contaminant in the porcine kidney cell line PK-15 (ATCC-CCL33). This agent was later shown to be a small, nonenveloped virus containing a single-stranded, circular DNA genome; it was named porcine circovirus (PCV). PCV antibodies in swine were found to be widespread, and experimental infections with this virus in pigs did not result in clinical disease, suggesting that PCV was nonpathogenic.
A new disease was described in Western Canada during the early and mid 1990s. The etiology was unknown, and the condition was named postweaning multisystemic wasting syndrome (PMWS). Affected pigs showed primarily poor growth rate, ill thrift, and/or wasting; histopathology identified characteristic systemic lymphoid lesions, among other lesions. In the late 1990s, an apparently novel PCV-like virus was isolated from PMWS-affected pigs. The new virus was antigenically and genetically distinct from PCV-1 and designated as PCV-2.
PCV-2 has been further associated with a number of disease syndromes in pigs, so the term porcine circovirus disease (PCVD) was proposed as a collective name. The terms PCV2-systemic disease (PCV2-SD) and PCV2-reproductive disease (PCV2-RD) have been proposed to replace PMWS and PCV2-associated reproductive failure, respectively. Moreover, it is currently thought that the most important PCVD is the PCV2-subclinical infection (PCV2-SI), which is linked with growth retardation without overt clinical signs. Porcine dermatitis and nephropathy syndrome (PDNS) is also included as a PCVD, although there is still no proof that PCV2 is the antigen linked with this immunocomplex disease.
PCV-3 has been recently linked with reproductive disorders and multisystemic inflammation associated with vasculitis, as well as with PDNS. The strongest evidence of disease association comes from in situ hybridization detection of viral genome associated with histopathologic lesions. Moreover, PCV-3 has also been found in a number of conditions (respiratory, digestive, neurologic), but its true association is still to be determined. Like the other porcine circoviruses, PCV-3 is found worldwide.
Etiology and Pathogenesis of Porcine Circovirus Diseases
Circoviruses are small (17–22 nm in diameter), nonenveloped viruses that contain a single strand of circular DNA. There are three types of porcine circoviruses, although only PCV-2 and probably PCV-3 are considered pathogenic. Phylogenetic studies have shown that at least four genotypes of PCV-2 exist (PCV-2a, b, c, and d). Novel genotypes have been found, and the most recent proposal suggests the existence of at least 8 PCV-2 genotypes. Some studies suggested a first genotype shift (from a to b) coincidental with major outbreaks of PCV2-SD in North America, Japan, and some European countries. In the past 5–7 years, another genotype shift (from b to d) has been evidenced epidemiologically. It is not clear whether differences in virulence exist among or within PCV-2 genotypes. So far, only one single genotype is considered to exist for PCV-1 and PCV-3.
Serologic surveys show that PCV-2 is widespread in swine, independent of the PCV-2-SD status of the farm. Results from retrospective serologic studies indicate that PCV-2 has been infecting pigs for >50 years so far, and phylogenetic studies indicate that PCV-2 has probably circulated in pigs during the past 100 years. PCV-3 is also widespread based on PCR results from many parts of the world, and it has been found to have existed in the 1990s.
Initially, PCV-2-SD was identified in high health herds that were free of most common swine pathogens. However, under field conditions, swine that show signs of PCV-2-SD usually are infected with multiple agents, including porcine parvovirus, porcine reproductive and respiratory syndrome virus, Mycoplasma hyopneumoniae, Actinobacillus pleuropneumoniae, Pasteurella multocida, Haemophilus parasuis, Staphylococcus spp, and Streptococcus spp ( See chapter Respiratory Diseases of Pigs). So far, the conditions by which PCV-3 is able to cause apparent disease are unknown. Moreover, the frequency of overt disease associated with this novel virus is also not known.
Accounts of multiple attempts to experimentally reproduce PCV-2-SD have been published. Some early trials (using tissue homogenates from pigs affected with PCV-2-SD or a PCV-2 isolate) reproduced PCV-2-SD–like histologic lesions but not the wasting condition. However, occasional studies subsequently reproduced clinical disease and lesions consistent with PCV-2-SD using only PCV-2, presumably, as inoculum. Consequently, it was suggested that PCV-2 infection, linked to other cofactors, was necessary for consistent development of full clinical disease. It appears that a number of factors, such as age and source of pigs, environmental conditions, genetics, the nature of the PCV-2 inoculum used, and the immunologic status of the pig at PCV2 infection, play a significant role in consistent experimental reproducibility of the disease.
The more consistent and repeatable PCV-2-SD disease models have been obtained using infectious and non-infectious cofactors as triggers. Also, the coinfection of PCV-2a and b genotypes has been linked to reproduction of clinical disease under experimental conditions. The mechanisms by which other viruses or immunostimulation may trigger the development of wasting in PCV-2-infected pigs is still unknown. High loads of PCV-2 in blood, lymphoid, and other tissues and in potential excretion routes are associated with expression of disease.
Damage to the immune system is the main feature of PCV-2-SD, suggesting that affected pigs have an acquired immunodeficiency. Lymphocyte depletion of lymphoid tissues, changes in peripheral blood mononuclear cell subpopulations, and altered cytokine expression patterns have all been demonstrated in pigs naturally and experimentally affected with the condition.
The identification of cells that support PCV-2 replication has been a matter of controversy. The large amount of PCV-2 antigen found in the macrophages and dendritic cells of diseased pigs appears to be the result of accumulation of viral particles. However, epithelial and endothelial cells seem to be the main target for PCV-2 replication, as well as a small proportion of macrophages and lymphocytes.
Much less is known regarding the pathogenesis of other PCVDs. PCV-2 is able to replicate in fetuses as well as in zona pellucida–free embryos. Moreover, an experiment with embryos exposed to PCV-2 and then transferred to receptor sows suggested that infection can lead to embryonic death. Therefore, it is believed that one of the potential outcomes of PCV-2 infection in sows could be return to estrus. Transplacental transmission of PCV-2 has been demonstrated. However, experiments using pregnant sows inoculated intranasally have yielded variable results. When successful, those studies have shown that PCV-2 may cause fetal death, similar to that of porcine parvovirosis, with live pigs together with dead piglets and mummies of different sizes. Late abortion has also been described in PCV-2 natural infection.
PDNS is considered a type III hypersensitivity reaction in which the antigen present in the immune complexes is unknown. It has been speculated that PCV-2 could be the antigen, but there is no definitive proof that PCV-2 causes PDNS lesions. Indirect evidence exists, such as significantly higher serum antibody titers to PCV-2 in affected pigs compared with healthy or PCV-2-SD-affected pigs. The association of PDNS with PCV-2 has also been suggested, but its causality is also yet to be demonstrated.
PCV-3 has not yet been isolated, and only one experimental infection (with an infectious clone) has been published in the peer-reviewed literature. This report claimed the reproduction of PDNS, but histopathologic- lesions were not convincing. No real data on the pathogenesis of PCV-3 infection exists as yet.
Epidemiology and Transmission of Porcine Circovirus Diseases
Porcine circoviruses are considered ubiquitous in countries with and without PCVDs. PCV-2 and PCV-3 infections have been described in wild boars; PCV-2-SD has also been described in wild boars. PCVDs associated with PCV-2 have been reported worldwide. Epidemiologic features are almost known only for PCV-2.
Transmission of PCV-2 may be by direct contact with infected pigs. PCV-2 has been detected in almost all potential excretion routes such as nasal, ocular, and bronchial secretions; saliva; urine; and feces. The virus can be found in semen, but the practical importance of this is probably negligible. Artificial insemination (AI) of sows with PCV-2-infected semen from experimentally inoculated boars did not result in sow infection or fetal infection. However, when such AI was performed with PCV-2-spiked semen, reproductive problems developed. Therefore, it seems that reproductive disease linked to AI is possible, but only when semen has a high virus load, which is unlikely under natural conditions. Although not demonstrated, it is assumed that contact with contaminated fomites, exposure to contaminated feeds or biologic products, multiple use of hypodermic needles, or biting insects may play a role in transmission.
PCV-2 may persist in swine for several months under either experimental or field conditions. Convalescent swine may carry virus for extended periods and be important in disease transmission. PCV-2 is fairly resistant to commonly used disinfectants and to irradiation, probably allowing it to accumulate in the environment and be infective for new groups of susceptible pigs if rigorous sanitary measures are not followed. The decline of colostral antibody titer in pigs is associated with onset of PCV-2-SD in late nursery or finishing pigs. Transplacental infection with PCV-2 has been documented, but it is not known whether pigs infected in utero are able to subsequently develop clinical PCV2-SD.
Some reports have suggested that animals other than swine may be infected with PCV-2 or PCV-3. However, results of serologic and PCR studies for antibody or genome against PCVs in other livestock species, non-suid wild animals, and pets have been contradictory, and experimental induction of disease using PCV-1 or PCV-2 in species of livestock other than swine has not been successful. Mice may be able to replicate and harbor PCV-2.
Clinical Findings of Porcine Circovirus Diseases
PCV-2-SD is characterized by overt weight loss. Disease often occurs in the fattening units in pigs 8–18 weeks old, although the disease can be also seen in older or younger pigs. Morbidity is typically 5%–20% among cohorts in the late nursery or finishing stages. Mortality in swine with signs of PCV-2-SD can occasionally be >50%. In addition to death loss, PCV-2-SD in finishing pigs may cause a substantial increase in time to reach market weight, resulting in economic loss. Growth retardation, wasting, and dyspnea are the clinical signs seen most frequently in outbreaks. Pallor, anemia, jaundice, diarrhea, and palpable inguinal lymphadenopathy also are seen in some affected pigs. A low-grade fever (104°–106°F [40°–41°C]) that lasts several days may be seen as well.
Overcrowding, poor air quality, insufficient air exchange, and commingled age groups seem to exacerbate the course of PCV-2-SD. Usually, only a few pigs in a group show wasting. The onset of disease may be acute, leading to death within a few days in some pigs. Other pigs show a more chronic disease and fail to gain weight or thrive.


Courtesy of Dr. Joaquim Segalés (Universitat Autònoma de Barcelona and Centre de Recerca en Sanitat Animal - IRTA).
PCV-2-SI is believed to occur in pigs that become infected with the virus and suffer from growth retardation (significantly lower average daily weight gain [ADWG]) but not overt clinical signs. In fact, in a farm affected by PCV-2-SD, a variable proportion of pigs developed the systemic disease, while most had only subclinical infection. PCV-2-SI had been unnoticed for many years until the advent of vaccines drew attention to this condition. Vaccinated pigs have an increased ADWG compared with non-vaccinated, apparently healthy counterparts. Such difference has been demonstrated to vary between 10−40 g/d, depending on the farm.
PCV2-RD characterized by late-term abortions and stillbirths in the absence or presence of other well-known reproductive pathogens seems to be the hallmark of clinical PCV-2 infection in sows. Most of these descriptions come from North America, usually in start-up herds. Return to estrus due to embryonic death as a potential outcome of intrauterine PCV-2 infection has been suggested based on experimental data. However, there are no field data to date unequivocally supporting this effect.
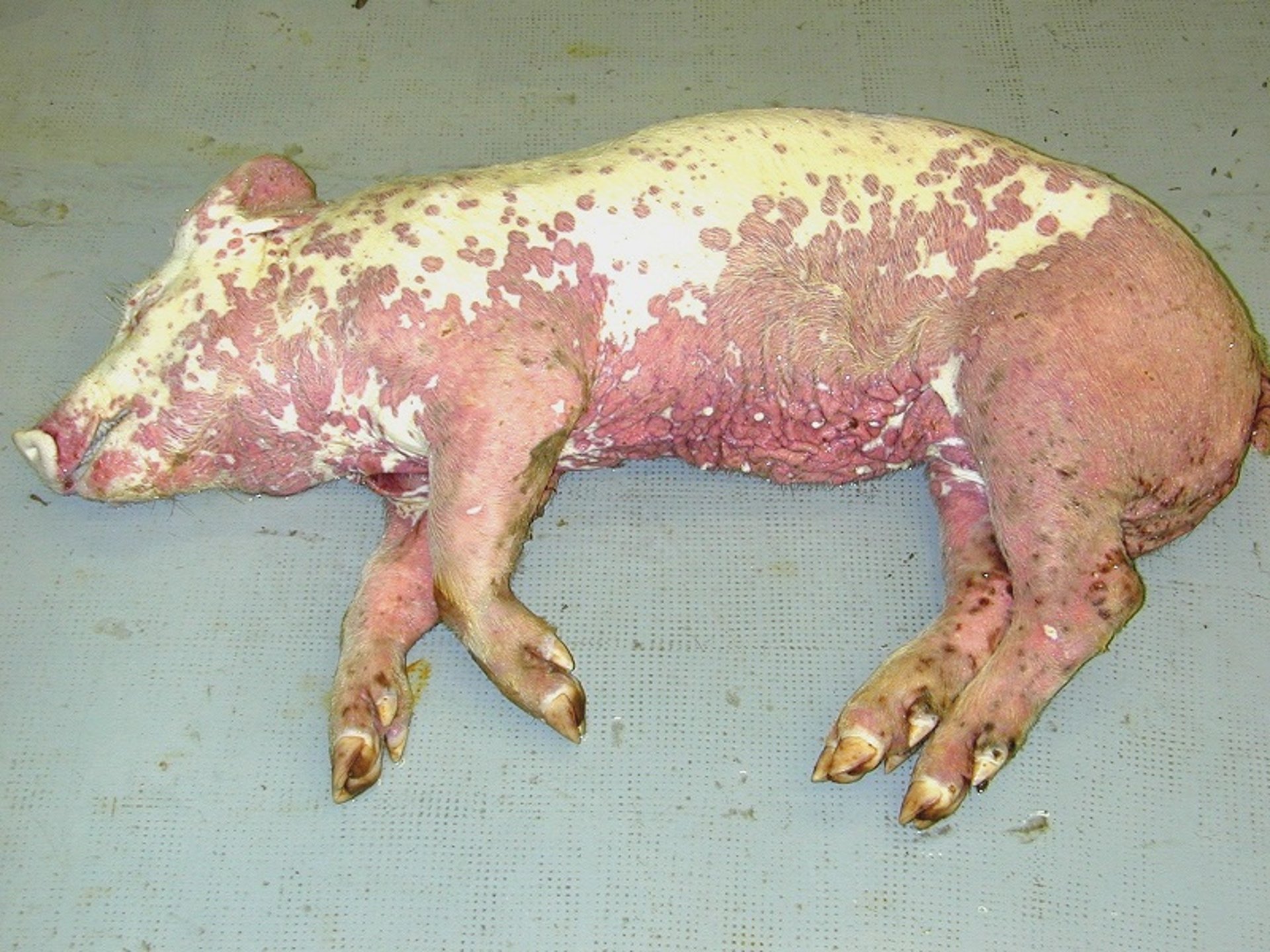
PDNS may affect nursery and growing pigs and, sporadically, adult animals. The prevalence of the syndrome in affected herds is relatively low (< 1%), although higher prevalences (>20%) have been described occasionally. Pigs with severe acute disease die within a few days after the onset of clinical signs, due to acute kidney failure with a significant increase in serum levels of creatinine and urea. Surviving pigs tend to recover and gain weight 7–10 days after the beginning of the syndrome. Affected pigs have anorexia, depression, prostration, stiff gait and/or reluctance to move, and normal temperatures or mild fever. The most obvious sign in the acute phase is the presence of irregular, red-to-purple macules and papules on the skin of the hindlimbs and perineal area, although distribution may be generalized in severely affected animals. With time, the lesions become covered by dark crusts and fade gradually (usually in 2–3 weeks), sometimes leaving scars. Although the most convincing evidence of association with PDNS is PCV-2 infection, this has never been experimentally demonstrated. In addition, PCV-3 has also been suggested to be linked with PDNS, but again, no demonstration of it has been gathered.
The PCV-3 genome has been found in many clinicopathologic conditions, including PDNS, reproductive disorders, respiratory disease, diarrhea, neurologic clinical outcomes, periweaning failure-to-thrive syndrome, and others. However, histopathologic data together with the detection of virus in tissues points to reproductive disease, multisystemic inflammation in postweaned pigs, and, with some contradictory results, PDNS.
Lesions
PCV-2-SD is grossly characterized by lymph nodes substantially enlarged and pale on cut surface, the thymus atrophied, and the tonsils thinner than normal. Splenic infarcts also may be present in a low proportion of pigs affected with PCV-2-SD. Histopathologic lymphoid lesions are characteristic, showing lymphocytic depletion and granulomatous inflammation, sometimes with the presence of multinucleate giant cells and amphophilic botryoid intracytoplasmic inclusion bodies of different sizes caused by accumulation of PCV-2 particles.
Lesions in the lung are common in affected pigs; their severity is influenced by duration of disease and presence of concurrent infections. Gross lung lesions may include failure to collapse, firmness, diffuse pulmonary edema, mottling, and consolidation. Microscopically, a variable degree of lymphohistiocytic interstitial pneumonia to granulomatous bronchointerstitial pneumonia with bronchiolitis and bronchiolar fibrosis can be seen.
Grossly, the liver may appear icteric and/or atrophic in a low proportion of affected pigs. Interlobular connective tissue may be prominent. Microscopic lesions range from single cell necrosis (apoptosis) with mild lymphocytic infiltration of portal zones to extensive lymphohistiocytic periportal hepatitis with diffuse necrosis of hepatocytes. The kidneys may be enlarged and show scattered to diffuse white foci on the cortical surface. Microscopic lesions include interstitial lymphohistiocytic infiltration. Other lesions seen in affected pigs include gastric ulceration (probably due in part to a prolonged fattening period in chronically affected pigs) and occasional multifocal lymphohistiocytic myocarditis. In severely affected pigs, lymphohistiocytic infiltrates can be seen in virtually all tissues.
PCV-2-SI pigs do not show gross lesions attributable to PCV-2 infection. These animals may show microscopic lymphoid lesions similar to those seen in pigs with PCV-2-SD, although only to a mild degree.
In PCV-2-RD, stillborn and non-viable neonatal piglets show chronic passive congestion of the liver and cardiac hypertrophy with multifocal areas of myocardial discoloration. The key histopathologic feature is fibrosing and/or necrotizing myocarditis in fetuses.

Courtesy of Dr. Joaquim Segalés (Universitat Autònoma de Barcelona and Centre de Recerca en Sanitat Animal - IRTA).
PDNS is easy to detect clinically because of the red-to-dark macules and papules, which correspond microscopically to necrosis and hemorrhage secondary to necrotizing vasculitis of dermal and hypodermal capillaries and arterioles. Necrotizing vasculitis is a systemic feature, but it is more prominent in the skin, renal pelvis, mesentery, and spleen (splenic infarcts may also be present as a result of necrotizing vasculitis of splenic arteries or arterioles).
Apart from skin lesions, pigs that die acutely with PDNS have firm, bilaterally enlarged kidneys, with a fine granular cortical surface and edema of the renal pelvis. The renal cortex displays multiple, small, reddish pinpoint lesions, similar to petechial hemorrhages, that microscopically correspond to enlarged and inflamed glomeruli (fibrinonecrotizing glomerulitis). Histologically, a moderate to severe nonpurulent interstitial nephritis with dilation of renal tubules is also seen. Usually, both skin and renal lesions are present, but in some cases, skin or renal lesions may occur alone. Lymph nodes may be enlarged and red due to blood drainage from affected zones with hemorrhages (mainly skin). Histopathologically, PCV-2-SD–like lesions such as lymphocyte depletion and histiocytic and/or multinucleate giant cell infiltration (although less severe) are usually found in lymphoid tissues of affected pigs, although to a milder degree.
The PCV-3 genome detected by in situ hybridization has been found in cases of reproductive failure (including weak-born neonatal piglets), PDNS, and acute death/wasting in 3–6-week-old piglets. Fetuses from reproductive disorders showed myocarditis, and weak-born neonatal piglets had encephalitis and myocarditis. Systemic periarteritis was the main feature of weaned piglets.
Diagnosis of Porcine Circovirus Diseases
Clinical signs
Histopathology
Serology

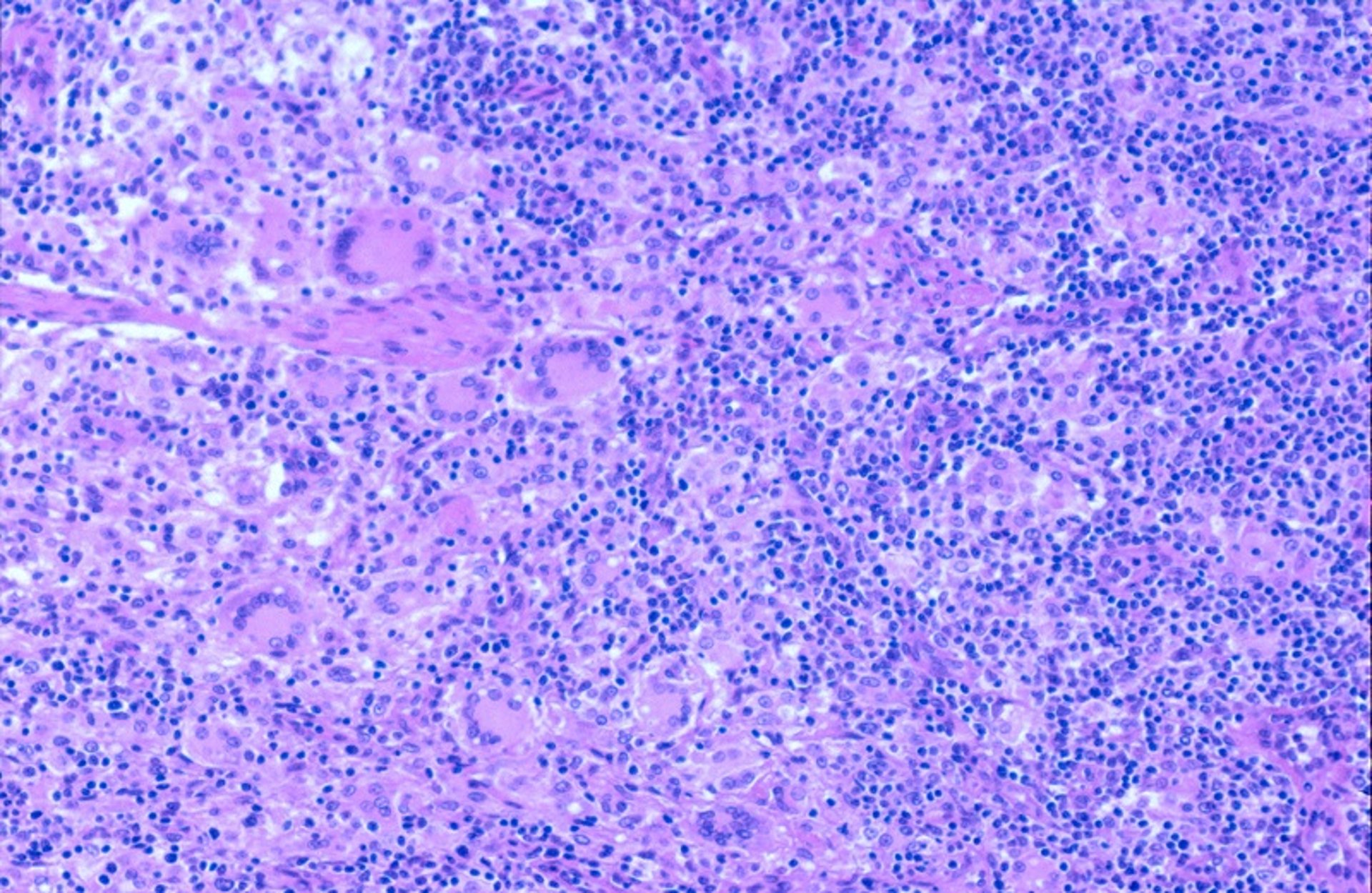
Courtesy of Dr. Joaquim Segalés (Universitat Autònoma de Barcelona and Centre de Recerca en Sanitat Animal - IRTA).
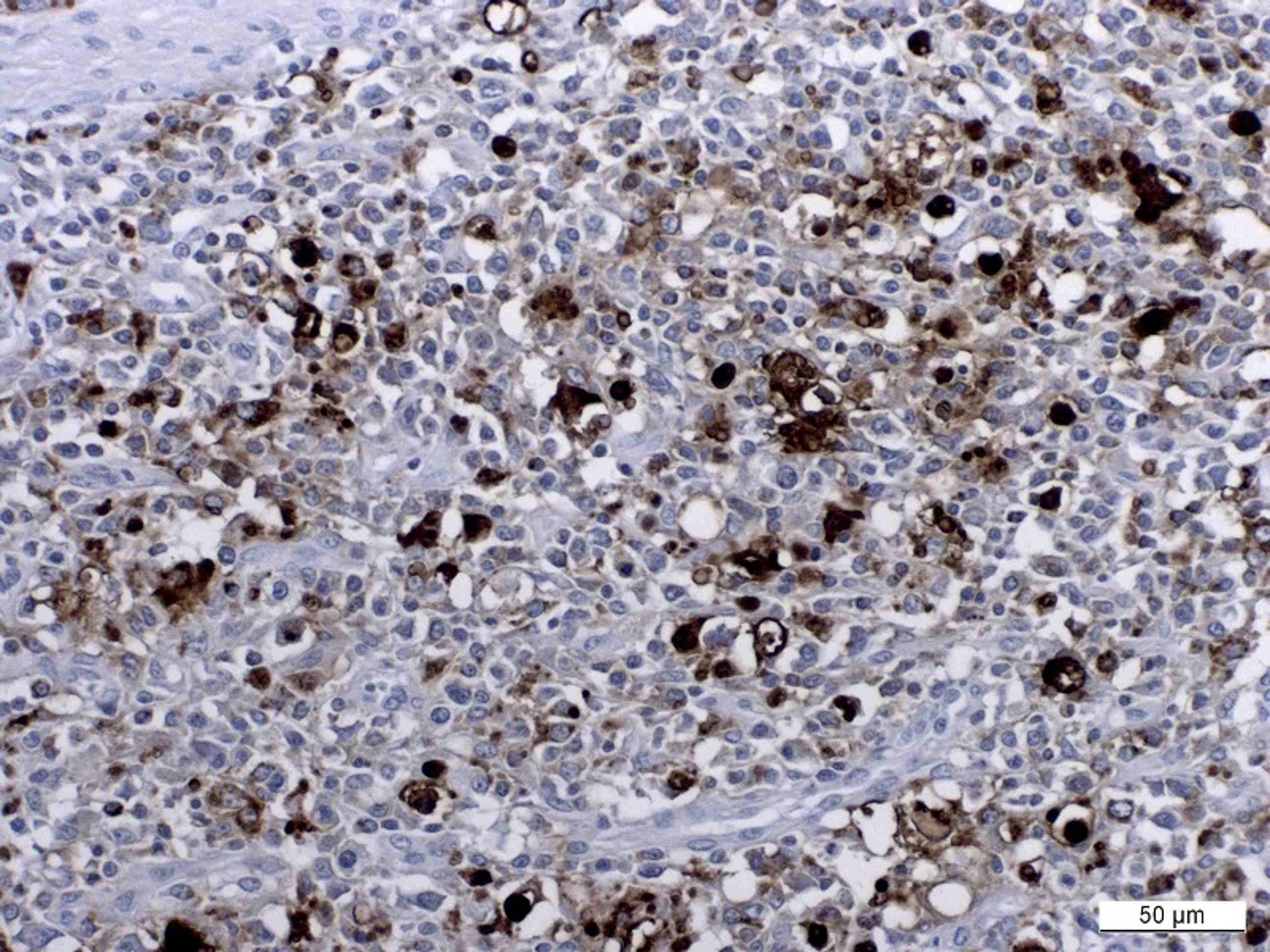
Courtesy of Dr. Joaquim Segalés (Universitat Autònoma de Barcelona and Centre de Recerca en Sanitat Animal - IRTA).
The porcine circovirus 2 systemic disease case definition includes three main diagnostic criteria:
clinical signs of wasting or ill thrift
presence of gross and microscopic (moderate and severe) lesions characteristic of the disease
presence of PCV-2 antigen or DNA (moderate to high amount) in the microscopic lymphoid lesions
Visualization of viral DNA or antigen in lesions is usually done using in situ hybridization or immunohistochemistry, respectively, and moderate to high amounts of virus are linked to the disease.
A herd case definition has been proposed, which includes two main criteria:
Significant increase of mortality and number of runt pigs or pigs failing to gain weight or thrive in comparison to previous values for the farm
Fulfillment of the three individual criteria listed above in at least one of five examined pigs
Differential diagnoses of PCV-2-SD include conditions causing increased mortality and growth retardation, such as:
and many others
Because PCV-2 is ubiquitous and the virus replicates in individual pigs for weeks to months, isolation of virus, detection of PCV-2 DNA in serum or tissues, or detection of PCV-2 antibodies in serum is not sufficient to establish a diagnosis of PCV-2-SD.
Antibodies against PCV-2 may be detected by ELISA (the most frequently used technique), indirect fluorescent antibody, or immunoperoxidase staining of infected cell cultures.
Viral isolation can be done on several porcine cell lines (mainly porcine kidney cells) using serum, bronchiolar lavage fluid, or tissue homogenates.
Viral DNA can be detected using PCR in most tissues or in serum from affected pigs. Several tissue samples from multiple pigs may be required to detect virus in cases of chronic disease. Virus quantification in serum by real time quantitative PCR (qRT-PCR) has been suggested as a potential diagnostic method in live pigs. Values of >107 PCV-2 genome copies/mL of serum usually have been linked with PCV-2-SD occurrence, although this value is test dependent and just an approximate epidemiologic assessment of the disease. However, PCV-2 infection is extremely common in clinically healthy pigs, and interpretation of positive qRT-PCR results is not always straightforward.

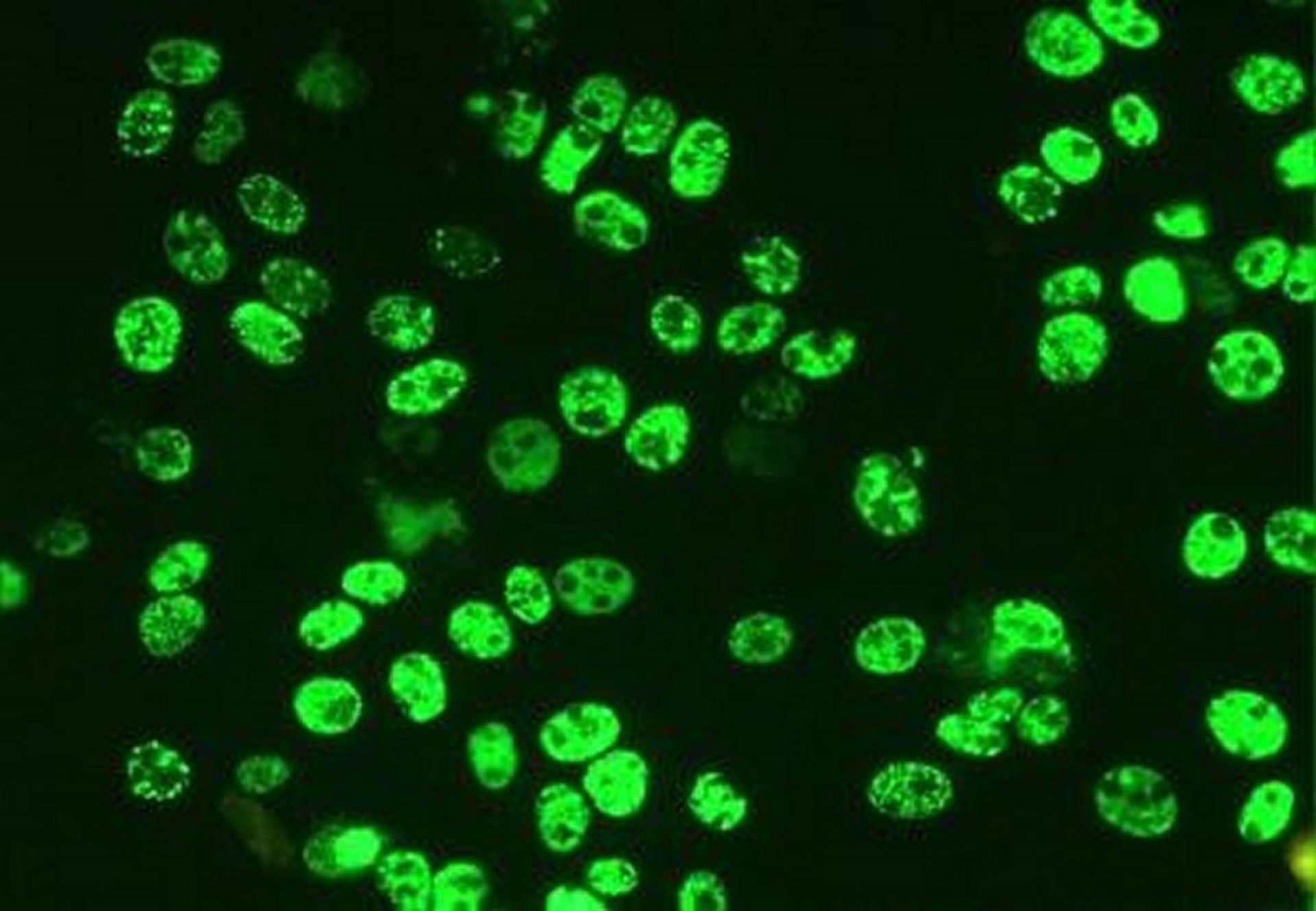
Courtesy of Dr. Gordon Allan.
The diagnostic approach of PCV-2-SI is of less interest for clinicians, because lack of overt clinical signs plus demonstration of infection by PCR would be enough to establish such a diagnosis. Values of < 105 or 106 PCV-2 genome copies/mL of serum usually have been linked with the subclinical infection.
The diagnosis of PCV-2-RD should include the following criteria:
late-term abortions and stillbirths, sometimes with hypertrophy of the fetal heart
extensive fibrosing and/or necrotizing myocarditis
high concentrations of PCV-2 in the myocardial lesions and other fetal tissues
Differential diagnoses for PCV-2-RD include:
pseudorabies (Aujeszky disease)
other diseases that cause late abortions, stillbirths, and weak piglets
If the effect of PCV-3 on reproduction is further confirmed, PCV-3 infection should also be included in the differential diagnostic list.
So far, there are no formal criteria to diagnose a putative return to estrus associated with PCV-2 infection. However, the occurrence of such signs together with evidence of viral circulation during the clinical episode should be demonstrated.
The case definition for PDNS is relatively simple and includes two main criteria:
presence of hemorrhagic and necrotizing skin lesions, mainly located on the hindlimbs and perineal area, and/or swollen and pale kidneys with generalized cortical petechiae
presence of systemic necrotizing vasculitis as well as necrotizing and fibrinous glomerulonephritis
Differential diagnosis of PDNS depends on the most significant pathologic outcome. Cutaneous manifestations may be confused with:
septicemic salmonellosis
porcine stress syndrome (malignant hyperthermia)
transit erythema (urine-soaked floors, chemical burns, etc)
other bacterial septicemias
Differential diagnoses for kidney lesions include:
Serum biochemical analyses may help differentiate PDNS from other diseases; urea and creatinine concentrations are markedly increased.
A PCV-3-associated disease definition is still lacking, and more data are needed to be sure this virus is truly pathogenic, causing disease in swine. Therefore, disease diagnostic criteria for PCV-3 infections are not yet defined. In contrast, different molecular methods to detect the viral genome (PCR and real-time quantitative PCR) have been developed and used in research or to monitor infection.
Treatment and Control of Porcine Circovirus Diseases
Vaccination
Because porcine circovirus 2 systemic disease is a multifactorial disease, effective control measures before the advent of PCV-2 vaccines were focused on control or eradication of these triggers. The most widely used control measures were: a) use of antibiotics to prevent concurrent bacterial infections; b) improvement of biosecurity and sanitary measures such as isolation of affected pigs and disinfection of pens after their use, decreasing stressors (eg, high stocking density, inadequate ventilation, inadequate temperature control); and c) control of concomitant viral infections, especially PRRS. Other prevention and control measures used on young pigs before the anticipated time of onset include injection of vitamins, IP injection of serum harvested from finishing pigs, and vaccination against common pathogens.
Currently, control of PCV-2-SD as well as PCV-2-SI is based on use of PCV-2 vaccines. There are four major commercial vaccines worldwide (plus a higher number with regional availability, mainly in southeast Asia). The first commercial vaccine was based on an inactivated PCV-2 isolate and was licensed for use in sows and gilts. The same vaccine was later licensed for use in piglets. Subsequently, three more vaccines have been developed, all for use in piglets ~2–3 weeks old or older. Two of these are subunit vaccines (PCV-2 capsid protein produced in a baculovirus system), and the third is an inactivated virus constructed by replacing the capsid gene of the nonpathogenic PCV-1 with that of PCV-2. In addition to significantly reducing mortality and runting percentages, these vaccines seem to improve ADWG, batch uniformity, slaughter weight uniformity, and feed conversion rate.
All major commercial PCV-2 vaccines are based on PCV-2a isolates, but cross-protection has been demonstrated against PCV-2b and PCV-2d. All PCV-2 vaccines are able to generate both cellular and humoral immune responses, which are believed to be the key features to control the subsequent PCV-2 infection that occurs under field conditions.
No treatment has proved successful for PDNS. Only those epizootic cases with moderate to high morbidity and mortality rates may be important in terms of economic losses. Treatment using a wide range of antimicrobial agents has been unsuccessful. Because the antigen responsible for triggering PDNS is not known, no preventive recommendations are indicated. Importantly, the use of PCV-2 vaccines worldwide has significantly reduced occurrence of this condition, emphasizing the putative implication of PCV-2 in its pathogenesis.
To date, no control measures have been specifically designed to control PCV-3 infections. Moreover, because the frequency of PCV-3 potentially associated disease is unknown, there is still no evidence for the need of a preventive product against this virus.
Key Points on Porcine Circovirus Diseases
Three types of porcine circoviruses have been described; PCV-2 is the most economically important one, and the impact of PCV-3 infection is unknown.
Major PCV-2 outcomes are PCV-2-SD (formerly PMWS), PCV-2-SI, PCV-2-reproductive disease, and PDNS; PCV-2-SD and PCV-2-SI are the most economically relevant.
PCV-2-SD shows moderate to severe lymphocyte depletion and granulomatous inflammation of lymphoid tissues; lymphohistiocytic inflammatory infiltrates can be found virtually in all tissues.
Diagnosis of PCV-2-SD is based on clinical signs, histopathology of lymphoid tissues, and PCV-2 detection in damaged tissues.
PCV-2 vaccination is the most cost-effective way to control PCVDs.
PCV-3 is apparently linked to reproductive problems and multisystemic periarteritis in weaned piglets.




