



Different subtypes of the avian metapneumovirus (AMPV A-D) have been isolated from commercial poultry and wild birds, which are considered natural reservoirs. These viruses were shown to not only induce respiratory disease (turkey rhinotracheitis, swollen head syndrome) but also reproductive disorders in different avian species. Molecular methods are used to detect and further characterize AMPV, although classic virus propagation from field samples is difficult. Currently, biosecurity in combination with vaccination is applied in the field to control infection and reduce the risk from secondary respiratory pathogens.
Avian metapneumovirus (AMPV) causes turkey rhinotracheitis (or avian pneumovirus infection of turkeys), an acute respiratory tract infection of turkeys. It is also associated with swollen head syndrome (or avian rhinotracheitis) in broilers and broiler breeders, as well as reproductive disorders, with a significant drop in egg production in chickens and ducks. The virus was first detected in turkeys in South Africa in the late 1970s and has spread to all the major poultry-producing areas in the world except for Australia. AMPV has been detected not only in chickens and turkeys but also in pheasants, Muscovy ducks, and guinea fowl. Geese, most other duck species, and possibly pigeons are suggested to be refractory to disease.
Epidemiologic studies provide evidence for the circulation of AMPV in wild birds, especially water-associated species. Some outbreaks have been attributed to vaccine-derived viruses, which may persist for several months in the environment. Infection with AMPV is often complicated by secondary bacterial infections, leading to high economic losses. In 2001, the first human metapneumovirus (HMPV) was isolated and classified as a member of the genus Metapneumovirus, which causes respiratory infections in people. Experimental studies suggest that turkeys also may be susceptible to HMPV. Complete genome sequencing confirmed that the genomic organization of HMPV is similar to that of AMPV. Overall, little is known about the cross-species pathogenicity of these two viruses.
Etiology of Avian Metapneumovirus
Avian metapneumovirus is a member of the family Paramyxoviridae, genus Metapneumovirus, which currently comprises the species AMPV and HMPV.
Isolates of AMPV are currently grouped in subtypes A to D. The sequence of the attachment glycoprotein (G protein) can be used to subtype different strains, because it is variable in length and sequence identity even within AMPV subtypes. Based on the phylogenetic analysis, it was suggested that the European subtypes A, B, and D are all more closely related to each other than to subtype C, which shows more similarity on the molecular level (sequence identity, genomic organization, codon usage bias, phylogenetic location) with HMPV. Whereas European and Asian AMPV C isolates can be grouped in one genetic sublineage, the other sublineage comprises the US isolates. The circulation of subtype D was described in France, but since 1980 has not been reported again.
Transmission and Epidemiology of Avian Metapneumovirus
Wild birds are considered natural reservoirs for avian metapneumovirus, and migratory birds may contribute to the distribution of the virus. A high apparent prevalence was recently determined particularly in mallards and American black ducks. The spread of AMPV appears to depend on the poultry population density, standard of hygiene, and biosecurity. Within or between poultry flocks, AMPV may spread rapidly horizontally by direct contact or by contact with contaminated material (morbidity rate up to 100%).
AMPV is assumed to be highly contagious. The enveloped virus is rapidly destroyed after release from the host to the environment. Because AMPV affects mainly ciliated epithelial cells of the upper respiratory tract, transmission is most likely to be airborne, especially by aerosol. Ciliated cells of the reproductive tract also may be target cells of AMPV. AMPV C was isolated from eggs of experimentally infected, SPF turkeys, but it was suggested that the vertical route may be short-lived and may play only a minor role in viral transmission.
Birds appear to shed AMPV for only a few days after infection. This short period of shedding suggests that there is no latency or carrier status under experimental conditions. There is evidence that, on farms, AMPV may persist for longer periods. Reconvalescent flocks may be repeatedly reinfected with AMPV within one fattening period.
Host Species, Pathogenesis, and Clinical Findings of Avian Metapneumovirus
Turkeys, chickens, and also ducks were shown to develop clinical signs with different subtypes of avian metapneumovirus. Whereas AMPV A and B are associated with clinical signs in chickens and turkeys, AMPV C of the North American lineage affects mainly turkeys, ducks of Asian and European lineage, and other commercial poultry less frequently, which may be subclinical infections.
Pheasants, Muscovy ducks, and guinea fowl kept in captivity may show signs, whereas geese, most other duck species, and pigeons are thought to be refractory to disease. Viral contact was also demonstrated for farmed ostriches. The American lineage of AMPV has been detected in sea gulls, sparrows, and additional wild bird species.
AMPV induces an acute, highly contagious infection of the upper respiratory tract. The main target cells are epithelial cells, but macrophages also may carry the virus. Infection leads to clumping and loss of cilia, which allows secondary pathogens to invade. Furthermore, studies indicate an immunosuppressive potential of the virus, which supports the replication of coinfecting pathogens. The clearance of AMPV coincides with the induction of AMPV antibodies and the disappearance of clinical signs.
Avian metapneumovirus affects all age groups, although younger birds seem to be more susceptible. In fattening turkeys the upper respiratory tract is predominantly affected, whereas in laying hens only a mild respiratory infection with a drop in egg production (up to 70%) and egg quality has been seen.
Coughing associated with lower respiratory tract involvement may lead to prolapse of the uterus in laying turkeys.


Courtesy of Dr. Rebecca Lindenwald.


Courtesy of Dr. Rebecca Lindenwald.
Typical respiratory signs of avian metapneumovirus in young turkeys include:
serous ocular and nasal discharge
frothy eyes
conjunctivitis
At later stages, signs include mucopurulent, turbid nasal discharge; plugged nostrils; swollen infraorbital sinuses; and snicking, sneezing, coughing, or tracheal rales. These respiratory signs are accompanied by depression, anorexia, and ruffled feathers.
The incubation period is 3–7 days. The mortality may be 1%–50% depending on age and constitution of the flock as well as secondary infections. Birds without secondary infections with good constitution may recover within 7–10 days. However, in birds with secondary infections and under poor management, the disease may be prolonged and exacerbated by airsacculitis, pericarditis, pneumonia, and perihepatitis.
Infection in chickens and pheasants is less clearly defined and may not always be associated with clinical signs. AMPV is associated with swollen head syndrome in chickens. This condition is characterized by swelling of the peri- and infraorbital sinuses, frothy eyes, nasal discharge, torticollis, and opisthotonos due to ear infection. Typically, < 4% of the flock is affected, although respiratory signs may be widespread. Mortality is rarely >2%. In broiler breeders and commercial layers, egg production and quality are frequently affected.
AMPV C infection of ducks may lead to respiratory signs and a drop in egg production (40%–85%) as well as poor egg shell quality. Clinically, signs may wane after 9–12 days if no secondary infections complicate the disease in ducks.
Although it was suggested that mice may be susceptible to AMPV C replication and develop lung lesions, other studies using another AMPV strain did not successfully establish an infection.
Lesions

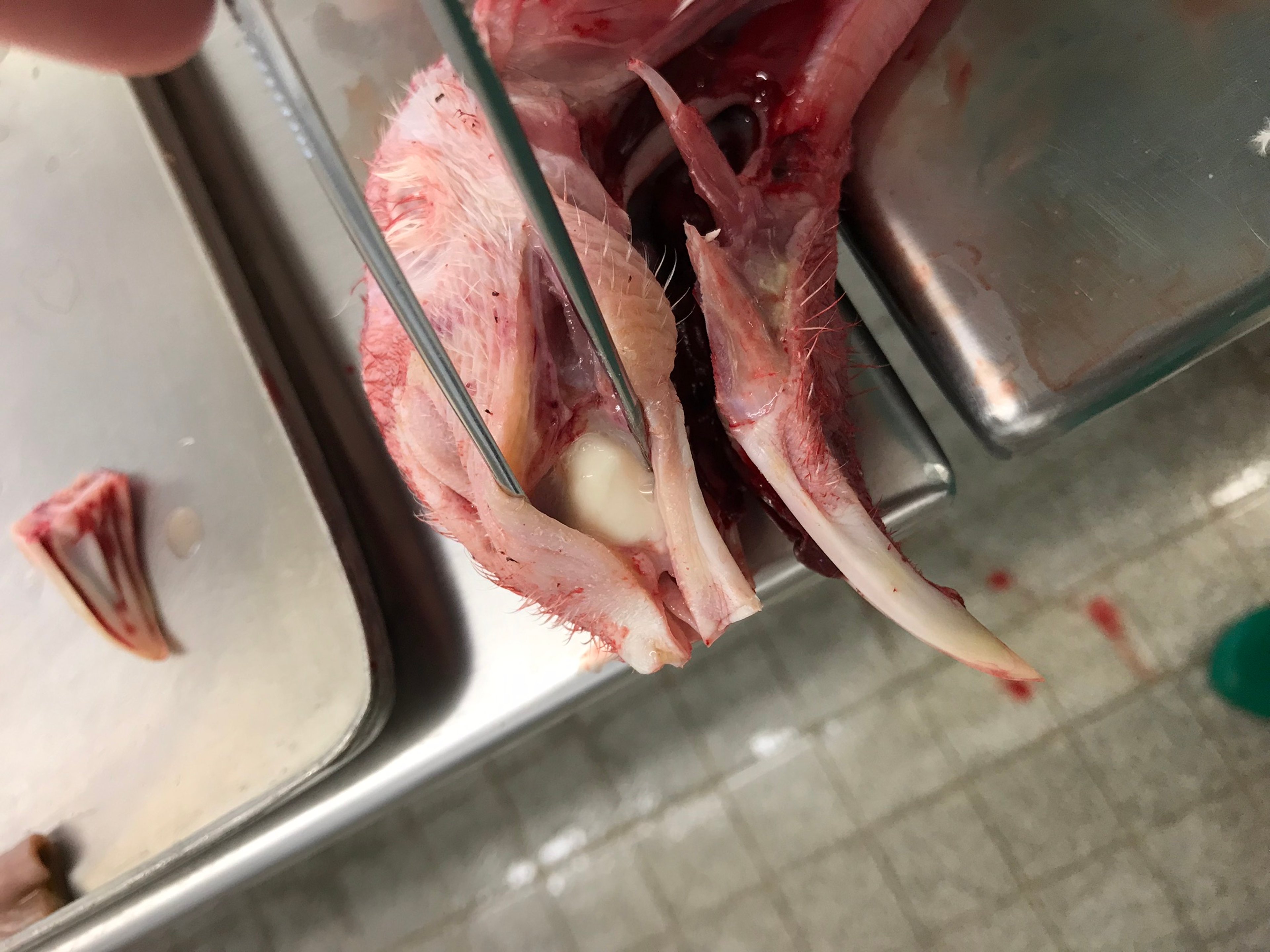
Courtesy of Dr. Arne Jung.
Macroscopic lesions depend on the course of infection, especially on secondary bacterial infections, and are most prominent on days 4–10 after infection. Gross lesions induced after experimental infection are due to rhinitis, tracheitis, sinusitis, and airsacculitis. Infected birds may be free of gross lesions. Serous to turbid mucus may be observed in the nasal cavity, nasal turbinates, trachea, and in infraorbital sinuses. During the course of infection, the secreted mucus turns from clear and serous to turbid and purulent.
Nonspecific signs of inflammation, such as swelling and hyperemia of the mucosa and excessive mucus, can be seen in the upper respiratory tract and in the air sacs. If secondary bacterial infections are present, copious inflammatory exudates are found in the respiratory tract. In addition, pneumonia, pericarditis, perihepatitis, splenomegaly, and hepatomegaly are seen. In the reproductive tract of laying turkeys, lesions can include egg peritonitis, ovary and oviduct regression, folded shell membranes in the oviduct, and misshapen eggs. Some infected birds are free of gross lesions.
Microscopic examination of the upper respiratory tract, including the secondary bronchi during the first 2 days after AMPV infection, reveals loss of cilia, increased glandular activity, congestion, and mild mononuclear infiltration of the submucosa. The most pronounced microscopic lesions are found in the mucosa or the nasal turbinates, which may be the most suitable tissue for microscopic evaluation. Harderian glands and lacrimal glands may also show infiltration of lymphocytes and formulation of lymphoid follicle-like structures in the interstitial tissue and around the secondary collecting ducts. The peak of microscopic lesion development in turkeys is expected 3–6 days after infection; it may be shorter in chickens.
Diagnosis of Avian Metapneumovirus
Virus detection and serology is necessary to diagnose avian metapneumovirus infection
Obtaining samples from the upper respiratory tract of birds in the early stages of the disease is extremely important when attempting avian metapneumovirus isolation. Especially in broiler-type chickens, samples should be taken before the sixth day after infection. Once clinical signs are obvious, the isolation of replicating AMPV may not be successful. The most suitable samples for AMPV detection are tracheal and choanal swabs. Tracheal organ cultures prepared from turkey or chicken embryos, or 1- to 2-day-old chicks, are the most sensitive for primary isolation of most AMPV subtypes. Ciliostasis may occur within 7 days of AMPV A and B but not subtype C inoculation or after passages.
The virus has also been isolated after the inoculation of 6- to 8-day-old embryonated chicken or turkey eggs via the yolk sac route and identified by electron microscopy, virus neutralization test, or molecular techniques. Cell cultures have not proved successful for the primary isolation of the virus. However, once the virus has been isolated and adapted in the systems above, it will grow in a variety of avian and mammalian cultures, inducing a cytopathic effect.
Reverse transcriptase PCR (RT-PCR), as well as real-time RT-PCR, tests targeting the F, N, or G gene of AMPV have been developed. Some systems are commercially available, and these techniques are widely used to detect virus in clinical material, particularly respiratory swabs. Samples may also be submitted on FTA cards for molecular diagnosis. Some nested RT-PCR tests have been constructed so that the subtype as well as the identity of virus can be determined from the clinical sample.
Based on the growing amount of genome sequence data and access to sequencing techniques, detailed characterization and molecular differentiation of isolated AMPV strains can be done, and vaccine and field strains may be identified. Thus far, sequencing approaches have not allowed a detection of host specific variations between strains. Multiplex systems allow the detection of a variety of co-infecting pathogens in addition to AMPV, as it was recently described using a nonfluidic PCR platform. The detection of new subtypes may require the development of new diagnostic tools.
Antigen detection tests have also been developed, including immunofluorescence and immunoperoxidase assays on both fixed and unfixed tissues.
Because of difficulties in isolation and identification of AMPV, serologic assays have been developed to confirm infection in commercial chickens and turkeys. A number of commercial ELISA kits are available and are commonly used, but other techniques, including virus neutralization and indirect immunofluorescence tests, have also been used. Both acute and convalescent serum samples should be submitted for analysis (paired serum samples from affected flocks). Although ELISA systems that use either subgroup A or B strains as antigens detect antibodies to both of these subgroups because of some cross-reactivity, the homologous antigen should be used for the efficient detection of subgroup C. The subtype specificity of the applied test may result in limited or no detection of other subtypes or new, emerging AMPV strains that do not cross-react.
Paramyxoviruses (particularly Newcastle disease, Avian Avulavirus-1 (AAV-1) and AAV-3), infectious bronchitis virus, and influenza viruses may cause respiratory disease and egg production problems in chickens and turkeys that closely resemble AMPV infection. These viruses can be differentiated on the basis of morphology, hemagglutinating and neuraminidase activity, and molecular characteristics. A wide range of bacteria and Mycoplasma spp can cause signs very similar to those of AMPV. These agents are frequently present as secondary opportunistic pathogens and may mask the presence of the AMPV.
Prevention and Treatment of Avian Metapneumovirus
Vaccination (live or inactivated) and improved biosecurity
Good management practices can significantly reduce the severity of avian metapneumovirus infection, especially in turkeys; in particular, optimal ventilation, stocking densities, temperature control, litter quality, and biosecurity have a positive influence on the outcome of the disease. The virus is sensitive to lipid solvents, stable at pH 3–9, and may be easily inactivated at temperatures above 50° C. Disinfectants such as quaternary ammonia, ethanol, iodophors, phenol derivatives, as well as bleach may be used to reduce the viability of AMPV. Some success in reducing disease severity by controlling secondary bacterial infections with antibiotics has also been reported.
Both live and inactivated vaccines are available for immunization of chickens and turkeys and are widely used in countries where the disease is endemic. Studies suggest that maternal antibodies may partially interfere with vaccine virus replication but overall do not provide sufficient protection against AMPV infection. A vaccination program should plan for the first immunization as soon as possible after hatching and may even be applied in the hatchery. It is crucial to achieve a homogenous state of immunization per flock and farm by application of an adequate vaccine dose to all birds.
Live vaccines, which may be applied by spray or drinking water in the field, stimulate both local respiratory and systemic immunity, and cross-protection between subtypes may occur. But live vaccines may induce only short-lived protection, especially for grow-out of toms, because of the fast decline of local immunity. Thus, repeated revaccination of turkeys is common practice. There is, however, a risk of reversion of the live vaccine strains to more virulent variants.
Inactivated AMPV vaccines are often used for booster immunization of layer and breeder flocks after priming with live vaccines. Whereas inactivated vaccines alone induce only partial protection against AMPV infection, the most efficient and long-lasting protection is achieved by a combined prime-boost vaccination program. This program comprises repeated priming with live-attenuated vaccines and booster immunization with inactivated adjuvanted vaccines. As experimentally shown, in ovo vaccination may also be a promising strategy for effective, early induction of an immune response. Besides live attenuated and classical inactivated vaccines, some genetically engineered viruses, including recombinant vectored vaccines, have been designed and tested under experimental conditions. These have induced partial protection and need further development.
Key Points
Avian metapneumovirus (AMPV) is present almost worldwide in commercial poultry flocks, and wild birds are considered natural reservoirs.
AMPV affects mainly the respiratory and reproductive tract, leading to respiratory signs and a drop in egg production, but significant economic losses are often also due to secondary bacterial infections.
A combination of hygiene measures and vaccination strategies contribute to the control of health problems and economic losses due to AMPV.




