

The liver is the largest discrete organ in the body.
The liver is uniquely supplied by arterial and venous blood. Oxygen-rich arterial blood provides ~20%–30% of hepatic perfusion, with the majority (70%–80%) supplied by the portal vein.
Blood flow to the liver in the proper hepatic artery provides ~25%–30% of liver perfusion and 75%–80% of its oxygenation. Arterial blood feeds into hepatic sinusoids and the peribiliary arterial plexus (PBAP). The PBAP provides nutrient vasculature, or vasa vasorum, for bile ductules and ducts and portal veins. In health, the hepatic artery maintains a perfusion pressure of ~70–90 mm Hg.
Blood flow to the liver from the portal vein (hepatopetal flow) traverses arborizing vasculature draining splanchnic viscera. This vasculature provides 70%–75% of total liver perfusion and delivers essential hepatotrophic substances (from the pancreas) that maintain normal hepatocyte and lobular size. The portal circulation wholly feeds into hepatic sinusoids, intermingling with the lesser arterial volume. As this venous system passively collects blood from a myriad of visceral capillaries, vascular resistance is high and blunts its perfusion pressure to ~5–8 mm Hg.
Whereas initial small portal vein tributaries have valves, the main trunk of the portal vein does not. Consequently, upstream resistance at any point along this system thwarts hepatopetal perfusion, detouring blood toward a path of lesser resistance (ie, via acquired portosystemic shunts [APSS]). This establishes hepatofugal portal flow (flow away from the liver).
In health, hepatic sinusoids have endothelium with numerous dynamic fenestrations. These permit sieving of ultralymph into the space of Disse (ie, space between the sinusoidal endothelium and microvillar hepatocyte surface). It is within this space that hepatic ultralymph exchanges nutrients and waste products (from the alimentary canal) with the liver, and accepts products synthesized or metabolized by hepatocytes for distribution within the systemic circulation.
Within sinusoids are Kupffer cells, the largest population of fixed macrophages in the body (~80%). These cells provide a crucial first line of defense against particulate debris, noxious products (bacterial toxins, endotoxin), and infectious agents translocated from the alimentary canal. Kupffer cells are endowed with a wide repertoire of metabolic capabilities; in addition to phagocytic scavenging, they also synthesize and release numerous inflammatory cytokines and mediators, designate first response to infectious agents and endotoxins, instigate initial immune-mediated responses and more chronic injury responses, and sequester iron. Macrophage iron accumulation heightens their reactivity; iron is accumulated from circulating free iron and senescent RBCs and released from heme moieties in P450 cytokines released from necrotic hepatocytes.
Within the space of Disse are stellate or Ito cells. When quiescent, these cells store retinoic acid (vitamin A) in a lipid-vacuolated phenotype. When activated by Kupffer cell mediators (ie, especially transforming growth factor B), Ito cells transform into a myofibroblast phenotype, the source of sinusoidal fibrillar collagen deposition within the space of Disse. These cells become the primary source of dissecting sinusoidal fibrosis in chronic necroinflammatory liver disease, a process sustained by inflammatory mediator release.
Because the portal vein transports blood directly from the splanchnic circulation (ie, blood draining from the intestines, colon, pancreas, and spleen), it delivers nutrients assimilated from food along with a myriad of pathogen-derived materials (ie, bacteria, bacterial toxins, lipopolysaccharide [LPS, also called endotoxin] from gram-negative bacteria, and other ingested toxins), along with antigens (ie, bacteria associated and food derived) translocated from the gut.
Inflammatory bowel disease (IBD) or other disorders disrupting the integrity of the intestinal or colonic barriers intensifies translocation of injurious molecules (ie, endotoxin, inflammatory cytokines, antigens) and bacteria to the liver. The high liver capacity for clearance of gut-origin molecules, pathogens, and antigens protects the body against wide systemic dispersal.
The high filtering surveillance capabilities of the liver reflect its large population of resident macrophages (hepatic Kupffer cells) and many other cells contributing to a broad repertoire of immunologic protection.
Among cells populating the normal liver, ~80% are hepatocytes, 3%–5% are cholangiocytes (biliary epithelium), ~10%–20% are liver sinusoidal endothelial cells (LSECs), ~4% are Kupffer cells, and ~3% are lymphocytes (estimates based on human and rodent liver studies).
Hepatic circulation is delivered through branches of the hepatic artery and the portal vein (see figure ).

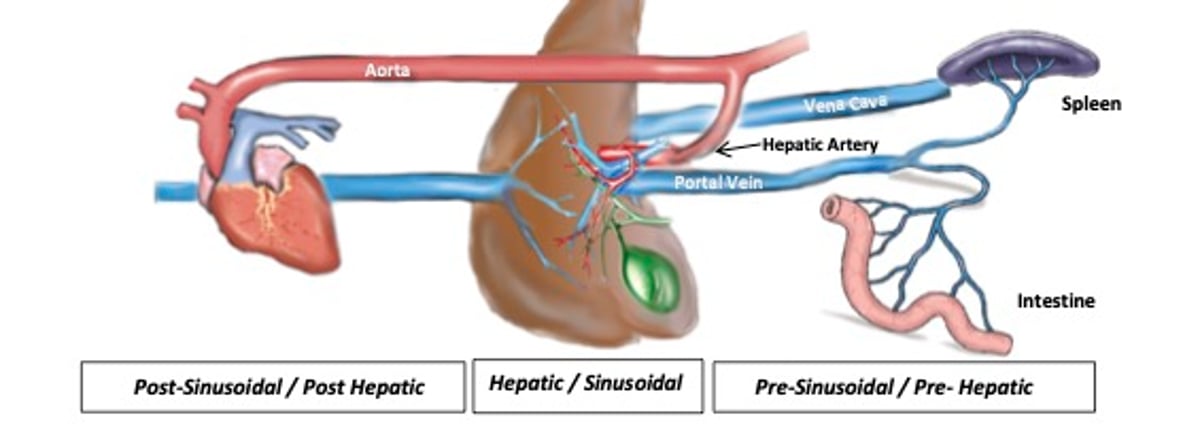
Schema of the liver, gallbladder and hepatic ducts, heart, spleen, and intestine as well as associated blood vessels. The hepatic artery, a terminal branch of the celiac artery arising from the abdominal aorta, is the major blood vessel that carries oxygenated blood to the liver. The caudal vena cava passes in the liver before reaching the diaphragm and terminating in the caudal part of the right atrium; hepatic tributaries embedded in the liver parenchyma drain into caudal vena cava as it courses through the liver. The portal vein brings functional blood to the liver from abdominal viscera such as the intestine and spleen. The gallbladder stores and concentrates bile—the exocrine product of liver cells. It can be useful to categorize hepatobiliary disorders (eg, ascites, jaundice, portal hypertension), on the basis of the localization of inciting factors or causes, as pre-hepatic (pre-sinusoidal), hepatic (sinusoidal), or post-hepatic (post-sinusoidal).
Courtesy of Dr. Sharon Center.
The functional organization of the liver is based on the , which can be divided into three metabolic zones (zone 1 = periportal, zone 2 = intermediate [trans-sinusoidal], zone 3 = centrilobular or periacinar) that designate a circulatory gradient originating from portal tracts and draining through hepatic venules and veins.

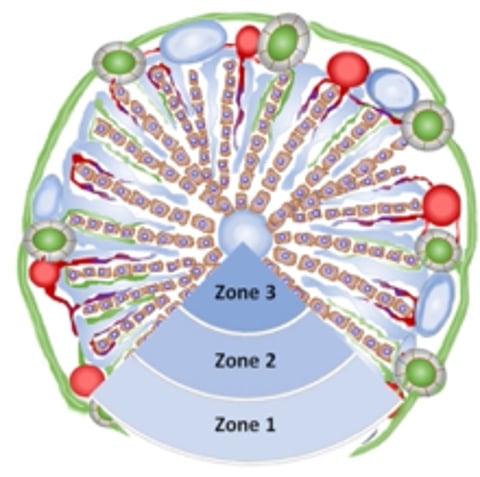
Schema of a classical hepatic lobule in cross-section illustrating zonal distribution: zone 1 = periportal, zone 2 = intermediate (trans-sinusoidal), zone 3 = centrilobular or periacinar.
Courtesy of Dr. Sharon Center.
Hepatic sinusoids are specialized microcapillaries transiting blood across the liver, bathing hepatocytes aligned in a linear radiating array of hepatic cords (mostly one cell in width). Sinusoids in zone 3 are orderly and linear, with less organization as cords transition to zone 1.
Sinusoidal architecture is supported by an extracellular matrix (ECM) scaffold that also provides intercellular structural cohesiveness. The ECM is composed of reticulin fibrils stained with reticulin silver stains. Reticulin fibrils represent variable combinations (depending on zonal or regional address) of proteoglycans, glycoproteins, and collagens.
In normal liver, types I and III fibrillar collagens prominently contribute to scaffolding of portal tracts and terminal hepatic venules and veins with only minor distribution along hepatic sinusoids. In contrast, sinusoidal ECM predominantly exhibits type IV network-type collagen. The proportional distribution of collagens, especially of those undergoing fibrogenesis, is remarkably altered in necroinflammatory liver disease.
Dual delivery of arterial and venous blood to portal tracts is accomplished through parallel ramifying tributaries of the hepatic artery and portal vein. These are not strictly dichotomous, in that one system may have fewer branches than the other at a given location. In health, ~25% of cardiac output circulates to the liver. Adjustment of transhepatic perfusion is complex but is principally influenced by sinusoidal tone under the influence of contractile perisinusoidal cells (stellate cells) and adjustments made directly by the LSECs.
In the dog, adjustments made by the throttling musculature surrounding medium and larger intrahepatic branches of the hepatic vein also influence transhepatic perfusion. Distribution of intrahepatic vasculature (ie, portal veins, hepatic arterioles) accompanies ductal tributaries of the biliary system.
Embryologically, portal tracts develop around the axis of the developing portal vein branches.
Bile formation initiates in hepatocytes where it collects into bile canaliculi (tiny intercellular membrane defined canals). These microscopic channels ramify into an expanding gradient of small (ductules) to larger ducts, ultimately delivering bile to the largest bile ducts, common bile duct, and gallbladder.

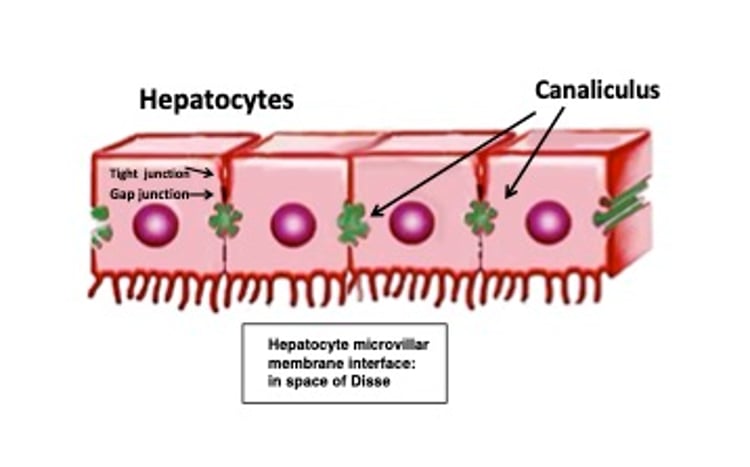
Simple diagrammatic representation of hepatocyte microvillar membrane interfacing with the space of Disse where they increase surface contact for exchange purposes (nutrients, toxins). Canaliculi, the first conduit of bile collection are specialized regions of hepatocyte membranes. Integrity is maintained by tight junctions whereas gap junctions facilitate intercellular exchange and passage of cells.
Adapted from Qayed E, Srinivasan S, Shahnavaz N, eds. Sleisinger and Fordtran's Gastrointestinal and Liver Disease Review and Assessment. 10th ed. Elsevier, 2016.
Hepatocytes are polyhedral cells exhibiting three specialized surface domains:
a sinusoidal border facing the perisinusoidal space of Disse
a lateral or intercellular domain segregating one hepatocyte from its adjacent neighboring hepatocyte
a specialized portion of the intercellular domain distinguishing the bile canaliculus, delimited by tight junctions (tightly adherent membrane regions)
The sinusoidal domain is covered by abundant microvilli facing the perisinusoidal space of Disse. These importantly increase the membrane surface area for absorptive and secretory exchange processes.
The vascular axis of portal tracts is the portal vein with its supportive ECM that provides scaffolding for the intrahepatic distribution of hepatic arterioles and biliary ductal elements. The ECM and adventitia (supportive connective tissue associated with vasculature) of portal regions render the hyperechoic appearance of portal tracts on ultrasonography.
Distributional branches of portal regions typically exhibit equivalently sized and duplicative profiles of hepatic arterial and biliary ductal elements, usually accompanied by a single portal vein profile. This structure defines the definition of portal triads (a triad of features: artery, ducts, portal vein).
Portal tracts distribute with a progressive diminishing size gradient from deep central liver lobe regions to peripheral subcapsular regions. In peripheral regions, diminutive portal tracts extend beyond the limits of the portal vein terminus, forming portal dyads that have only a tiny arterial and ductile profile (no portal vein). Considering all portal tracts distributed across the liver, 70%–80% are in triad configuration and 20%–30% are in a dyad configuration, with an occasional rare single arterial profile (referred to as an orphaned arteriole).
A commonly used anatomic landmark is the limiting plate of the portal tract (see figure ). This is the area where the portal tract ECM intersects with the limiting plate of hepatic cords (hepatocytes) defining the portal tract margin. This landmark is used for defining whether inflammatory cells invading portal tracts qualify as causing zone 1 (or portal) hepatitis. If this margin is unequivocally disrupted in numerous portal tracts, with inflammatory cells intermingling with hepatocytes with or without focal hepatocellular injury, a diagnosis of portal hepatitis is qualified.
Liver sinusoidal endothelial cells (LSECs) are delicate cells lining the low shear hepatic sinusoidal capillaries. Despite representing the most abundant nonparenchymal cells in the liver (~15%–20% of liver cells), LSECs constitute only ~3% of liver mass. Functioning as a first-line filtration barrier between circulating blood and the sinusoidal domain of hepatocytes, these cells additionally participate in critical physiologic and immunologic functions.
The unique structure of LSECs distinguish the sinusoidal capillaries as the most permeable endothelial surface in the mammalian body. Unlike microcapillaries elsewhere, hepatic sinusoids lack a continuous basement membrane and exhibit large dynamic fenestra (holes). Their permeable structure facilitates rapid collection of a plasma ultrafiltrate (lymph) in the space of Disse.
The LSECs provide vital metabolic, secretory, storage, clearance, surveillance, scavenging, and immunological functions. These cells demonstrate one of the highest endocytic capacities in the body, a function that orchestrates clearance of soluble immune complexes, albumin, and numerous circulating wastes.
LSECs also synthesize and store components of the ECM, synthesize and metabolize inflammatory mediators (including but not limited to nitrous oxide, endothelin, prostaglandins, cytokines, and chemokines), and engage in cross talk with other immunoreactive cells. Consequently, LSECs substantially influence innate and adaptive immune responses within the liver. Furthermore, the dual surveillance afforded by Kupffer and LSECs in the liver provides the most commanding scavenger network in the body.
Surveillance activity of LSEC is optimized by collaborative cell features:
unique sinusoidal porosity
moderated expression of pathogen-associated molecular pattern (PAMP) receptors (eg, toll-like receptor 4 that mitigates pathological endotoxin/LPS exposure)
presence of promiscuous scavenger receptors
dynamic endocytic capacity
capability for recruiting leukocytes to areas of injury and inflammation
function as antigen-presenting cells important for lymphocyte antigen sensitization.
Considering the myriad functions and interactions of LSEC and their essential role in sinusoidal perfusion, it is easy to understand why sinusoidal fibrosis disturbs liver function in complex ways.
The space of Disse, situated between LSECs and the sinusoidal microvillar surface of hepatocytes, is an extensive and unique extravascular space. Ultrafiltrate formed from sinusoidal blood in this space facilitates hepatocellular exchange processes and represents hepatic lymph. Distribution of the ultrafiltrate remaining in the space of Disse follows one of four possible routes:
to portal lymphatics within an areas with a loose adventitial collecting network within the space of Mall
to a loosely woven adventitial lymph collecting network surrounding branches of the hepatic vein in centrilobular regions (zone 3)
to a loosely structured adventitial lymph collection regions beneath the liver capsule (Glissen’s capsule), this fluid transiting paracellular gaps across hepatic cords
to a reentry into sinusoidal blood through its highly porous structure
Ultrafiltrate achieving lymph channel collection flows to regional lymph nodes, gradually making its way into the cisterna chyle and thoracic duct to reenter the systemic circulation. Disorders causing intrahepatic sinusoidal hypertension (see section on ascites formation) can lead to a “weeping” of ultrafiltrate from the liver capsule into the abdominal cavity and, rarely, the thorax.
In health, the ECM in the space of Disse is predominantly produced by hepatic stellate cells, with minor contributions from the LSECs. This material constitutes the reticulin substructure supporting and tethering sinusoidal and lobular architecture, morphologically assessed in liver biopsies with application of a reticulin stain (silver stain). Composition and density of the ECM importantly influences the biology of hepatocytes, sinusoidal endothelium, stellate cells, and the normal zonal heterogeneity of parenchymal and nonparenchymal cell populations across the liver.
Functional heterogeneity of various hepatic cell populations influences liver structure and function and is highly sensitive to altered sinusoidal perfusion, oxygenation, and nutrient dispersal. The normal ECM nearest centrilobular regions is rich in fibronectin, type III collagen. and dermatan sulfate, whereas the ECM nearest to portal regions is rich in laminin, type IV collagen, and heparin sulfate.
Disturbance in the compositional structure, sinusoidal perfusion, architectural remodeling, and fibrosis wrought by liver injury, importantly, influence cell functions across the liver. In some cases, drift in cell phenotypes and functional heterogeneity rather than loss of hepatic mass underlie important hepatic functional disturbances.
Hepatic stellate cells have historically been referenced as Ito cells, hepatic lipocytes, fat-storing cells, and parasinusoidal cells. By consensus, the preferred nomenclature is hepatic stellate cell. These cells are irregularly distributed in the space of Disse and have a broad functional repertoire and cell plasticity. Among their numerous functions are:
retinoid (vitamin A) storage and homeostasis
ECM deposition and remodeling (elaborating matrix components and matrix metalloproteinases, phagocytosis of apoptotic cellular debris)
response to a range of pathogen-associated molecular patterns (PAMPs)
regenerative influence (developmental and in response to injury) through elaboration of growth factor and cytokines
regulatory influence on sinusoidal vascular tone (angiotensin and neuroresponsive receptors)
involvement in intermediary metabolism and response to xenobiotics (drugs, toxins)
immunoregulatory functions (induction of immune responses and antigen tolerance)
Stellate cells comprise approximately one-third of the nonparenchymal cells in the liver and ~10%–15% of the total resident cells. Morphologically, stellate cells have long cytoplasmic processes encircling hepatic sinusoids. A single stellate cell usually surrounds or interfaces with at least two nearby sinusoids. As such, these cells can noticeably influence sinusoidal tone and perfusion.
Whereas other mesenchymal cell populations in the liver can contribute to ECM accrual and remodeling, stellate cells play a dominant role. Activated stellate cells play an overriding role in development of perisinusoidal fibrosis characterized by accumulation of types I and III collagens in the space of Disse. Such deposition appreciably impairs ultrafiltrate formation and normal hepatocyte-to-ultrafiltrate/sinusoidal exchange processes essential to normal liver function.
Upon activation, a stellate cell morphologically transforms from a vacuolated vitamin A–storing cell to one that is fibrogenic, exhibiting a contractile myofibroblastic phenotype (expressing alpha-SMA [alpha-smooth muscle actin]). Yet stellate cells are not primarily involved in the early stages of periductal fibrosis exhibited in cholangitis, cholangiohepatitis, or extrahepatic bile duct obstruction (EHBDO). Rather, a small resident population of cells surrounding the portal vein, which normally manage the integrity of the portal tract, proliferate and differentiate into myofibroblast-like cells producing type 1 collagen.
Hepatic Kupffer cells are large intravascular macrophages situated against the sinusoidal endothelium (see figure ). These cells represent 90% of the fixed macrophages in the body and ~35% of the nonparenchymal liver cell mass. Virtually every hepatocyte is close to a Kupffer cell. While regarded as fixed macrophages, these cells locally migrate along hepatic sinusoids.
Kupffer cells are opportunistically positioned to influence hepatic homeostasis, tolerogenic and reactive immune responses, and the initiation and progression of liver disease. These cells influence liver homeostasis through mediation of immune response against nonpathogenic materials (phagocytic removal and tolerance), participation in coordinated responses targeting pathogen clearance, leukocyte recruitment, and antigen presentation to lymphocytes (eliciting either tolerogenic or immunoreactive response).
The primary function of Kupffer cells is to discriminate and remove particulate and soluble materials from the sinusoidal circulation. Targeted materials include translocated enteric bacteria or bacteria-related debris, endotoxin, degenerate host cellular debris (eg, senescent RBCs), circulating tumor cells, and a variety of macromolecules including immune complexes.
Limiting endotoxin exposure involves a tolerogenic adaptation to the near-constant endotoxin exposure from the portal circulation. Without this adaptive tolerance, endotoxin would continuously incite toll-like receptor-4 (TLR-4) expression that can initiate an inflammatory response. As noted previously, the dual presence of Kupffer cells and LSECs in the liver provide essential surveillance functions. This activity and the phagocytic capacity of Kupffer cells is substantially impaired by hepatic architectural remodeling and fibrosis that disrupt sinusoidal circulation and by many forms of cholestatic liver disease (especially EHBDO).
Antigen presentation to lymphocytes by Kupffer cells, in concert with LSECs and circulating dendritic cells, can initiate hepatic inflammation. Activated Kupffer cells exhibit pathological TLR-4 expression and elaborate IL-2, tumor necrosis factor (TNF)-alpha, and IL-10 that initiate or augment an inflammatory response. These cells also can induce cytotoxic lymphocytes often involved in or evolved from chronic hepatitis.
With pathological activation, Kupffer cells enlarge and display phagocytic activity that can be observed in routine liver biopsy sections. In some circumstances, Kupffer cell enlargement seemingly can disrupt the normal rheology of blood flow across hepatic sinusoids. Kupffer cell activation is common in animals with inflammatory bowel disease, in which a milieu of bacteria, bacterial products including endotoxin, and particulate and soluble debris undergo enhanced enteric translocation.





