



Canaliculi are the first conduits transporting hepatocyte bile (diameter, ~1 mcm). The canalicular membranes represent a specialized component of the hepatocyte cell membrane delimited by tight junctions; these segregate bile from sinusoidal blood and the ultrafiltrate accumulated in the space of Disse. A web of contractile pericanalicular microfilaments propel bile from canaliculi into cholangioles (first-level biliary ductular element).

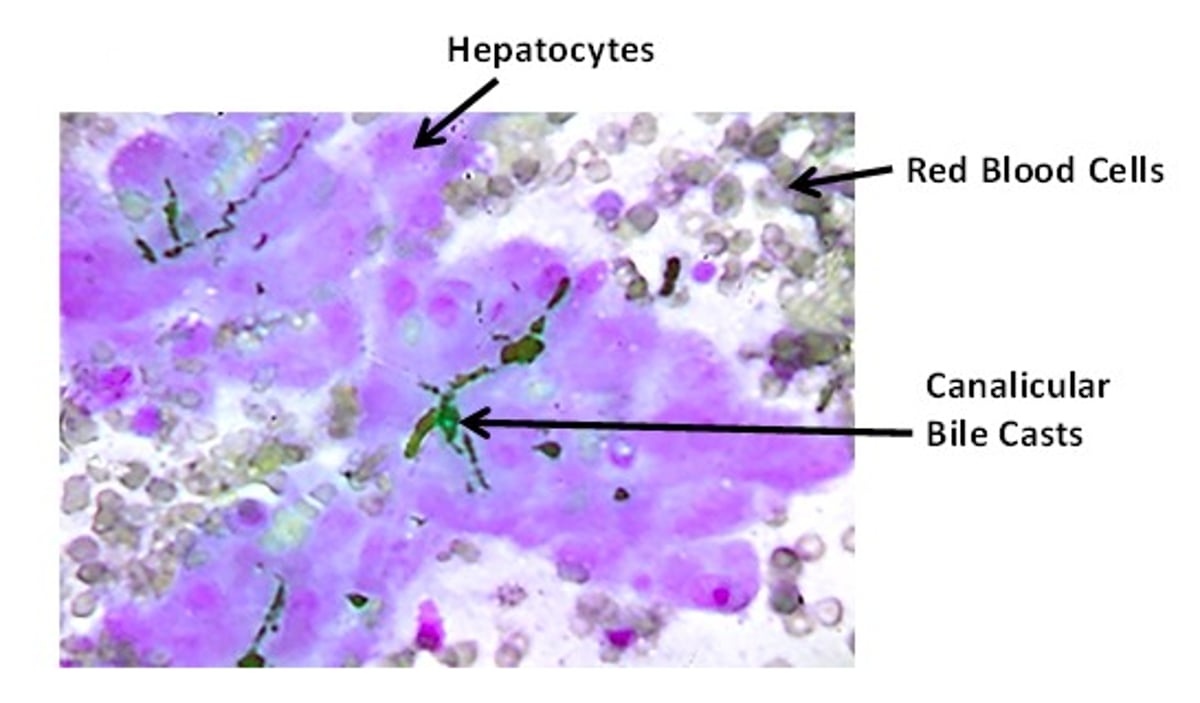
Cytologic aspirate demonstrating dark green-black canalicular casts between adjacent hepatocytes. Red blood cells stain gray-purple. This sample was collected from a dog with gram-negative sepsis that was jaundiced with no elevation in liver enzyme activity. Modified Wright-Giemsa stain; original magnification, 400×.
Courtesy of Dr. Sharon Center.

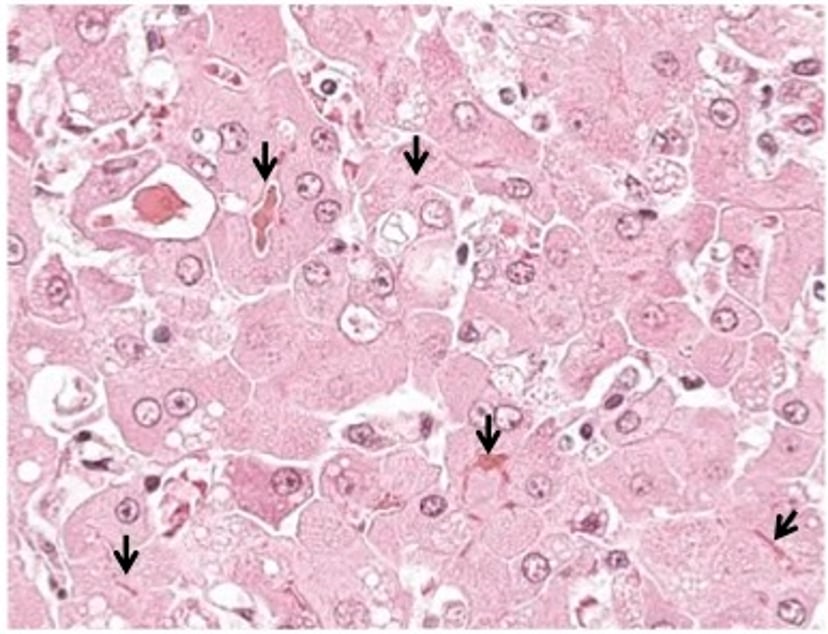
Histologic section from a dog with critical necroinflammatory liver disease (hepatitis) demonstrating severe canalicular bile casts (arrows). This dog was jaundiced with markedly increased ALT, AST, alkaline phosphatase (ALP), and gamma glutamyltransferase (GGT) activities. H&E stain; original magnification, 1000×.
Courtesy of Dr. Sharon Center.
The term biliary tree or biliary tract denotes the interconnected biliary pathways that transport bile from hepatocytes to the common bile duct and gallbladder. Bile initially collects into canaliculi of hepatocytes and then is transported retrograde relative to intrahepatic blood flow (bile flows from centrilobular regions toward portal tracts; blood flows from portal tracts to centrilobular regions) through cholangioles to small to medium-sized interlobular bile ducts, and then converge into progressively larger bile duct elements.

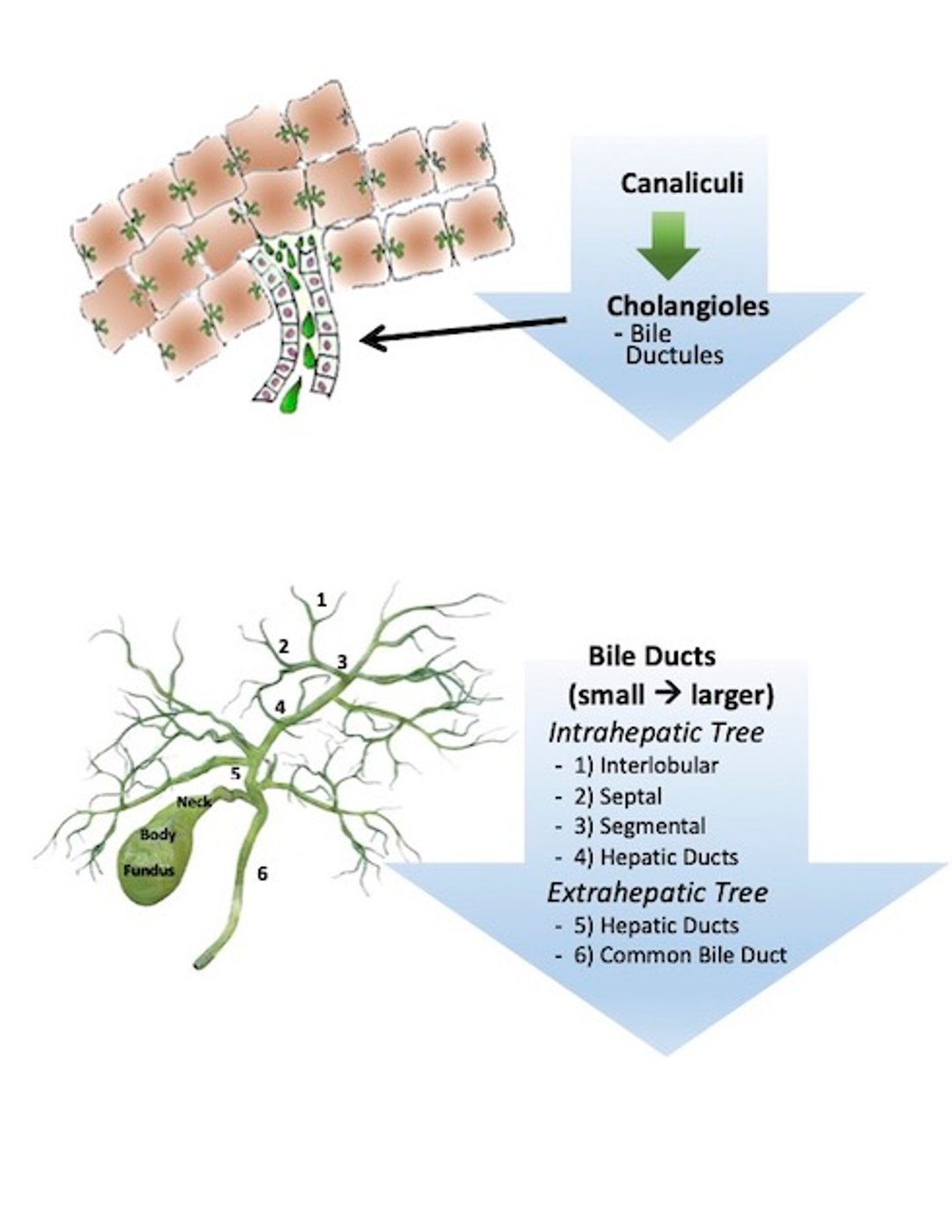
Diagrammatic illustration of microscopic canaliculi, the first components of the intrahepatic biliary tree. Bile flows from canaliculi into cholangioles (minute bile ductules lined by cuboidal relative undifferentiated biliary epithelium that have a basement membrane but no designated capillary). These merge into interlobular bile ducts, accompanied by a branch of the hepatic artery and portal vein, and then into progressively larger ducts until hepatic ducts form.
Courtesy of Dr. Sharon Center.
Canalicular bile first flows through canals of Hering (primitive cholangiocytes with canaliculi on one side and a cholangiole or bile ductule on the other). Cholangioles drain liver lobules and traverse portal tracts without blood vessels. These form an interconnected branching system of minute ductules, partly lined by hepatocytes and relatively undifferentiated cholangiocytes (biliary epithelial cells); they have a basement membrane but no distinct capillary supply.
Cholangioles converge, forming interlobular bile ducts, the smallest branches of the intrahepatic biliary tree composed of cuboidal epithelium and accompanied by vasculature (branches of hepatic artery and portal vein). Convergence of two or more interlobular bile ducts form larger septal bile ducts, lined by cylindrical cuboidal epithelium. These converge forming larger segmental bile ducts, which converge into hepatic ducts (minimal in dogs and cats). Categorical naming of ducts is based on size and epithelial morphology (flattened, cuboidal, cylindrical, columnar).
The extrahepatic biliary tree includes the hepatic ducts, common bile duct, and the gallbladder. The gallbladder is attached to the liver, between the right medial, left medial, and quadrate liver lobes, at the gallbladder fossa by loose connective tissue (small veins, lymphatics). Cholecystecomy (surgical removal of the gallbladder) requires blunt dissection of this area to gently detach this region before the gallbladder can be removed.
Liver tissue adjacent to the largest intrahepatic bile ducts and near the gallbladder fossa exhibits periductal glandular elements (intramural and extramural peribiliary glands). Extramural peribiliary glands (seromucinous glands) display a complex branched tubule-alveolar structure that can be confused with a proliferative-type DPM, whereas intramural peribiliary glands can be confused with a Caroli-phenotype DPM.
These structures are most often encountered in liver biopsies that have been collected near the gallbladder fossa (parenchyma with adventitial and ECM attachments to the gallbladder) at the time of cholecystectomy. These areas should not be submitted as liver biopsy samples as they are not representative of other portions of the liver and may lead to an inaccurate diagnosis.
Relatively undifferentiated biliary epithelial cells (cholangiocytes) represent a minor subset of cells in the liver (~3%–5%). Yet these cells are essential to bile production and flow and for the compositional modification of bile. Approximately 70% of bile flow is generated at the canalicular level, with ~30% attributed to cholangiocytes.
Regulation of bile volume and flow is complex, involving numerous membrane transporters, exchangers, and specialized ion channels. During transport in the biliary tree, bile is modified by cholangiocytes (eg, secretion or absorption of electrolytes, glucose, amino acids, water, and other organic solutes).
Once considered quiescent cells assigned predominantly to bile management, cholangiocytes are now understood as dynamic metabolically active cells. Wide morphological and functional diversity exists among cholangiocytes across different segments of the biliary tree.
Cell differentiation generally coordinates with ductule or duct size. The smallest ducts (cholangioles) are populated by the least differentiated cells. Cholangiocytes acquire an activated or reactive phenotype integrally involved in the inflammatory milieu of necroinflammatory liver disease, whether or not bile ducts are directly targeted.

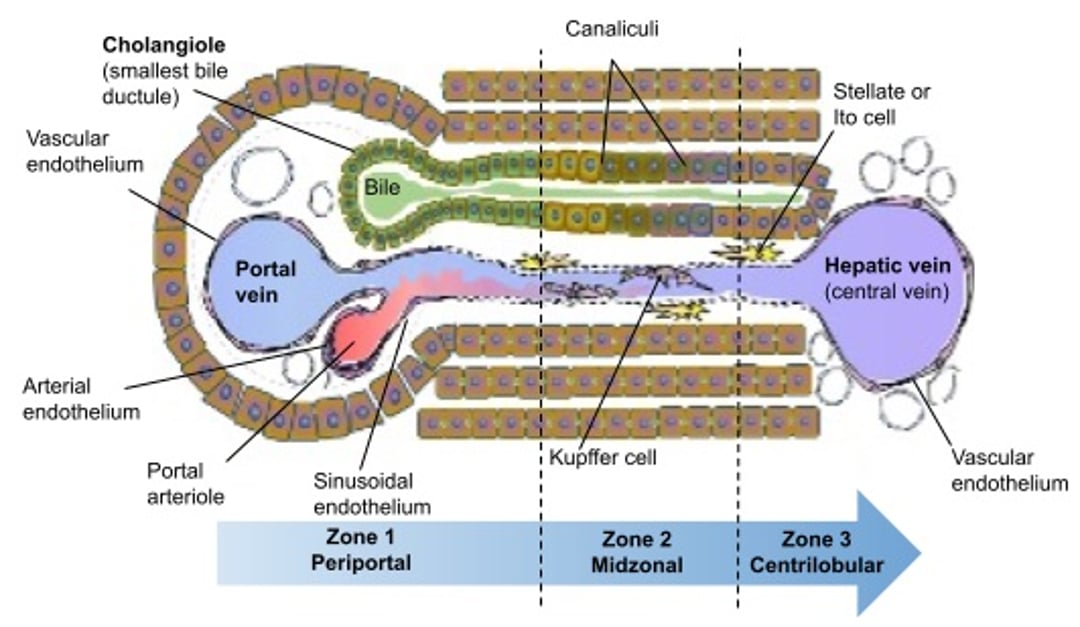
Schema of a hepatic acinus (the functional subunit of a hepatic lobule) in cross-section illustrating blood and bile flow relative to the portal tract (zone 1)—in this case a portal triad—and centrilobular (zone 3) regions. The portal field (containing portal venule, bile ductule and hepatic arteriole) is connected to the central vein by sinusoids. Hepatocytes, endothelial cells and many other cells types are exposed to blood flowing centripedally from the portal to the central area. Bile flows centrifugally towards the portal triad. In the sinusoids, nutrient-rich blood from the portal venule mixes with oxygen rich blood from the hepatic arteriole. Displayed are the orientation of hepatocytes with their canalicular conduits (green intercellular structures), single and occasionally double-wide hepatic cords, the smallest biliary ductular element (cholangiole), sinusoidal endothelium, intrasinusoidal Kupffer cells, and extrasinusoidal stellate cells (within the space of Disse).
Courtesy of Sharon Center.

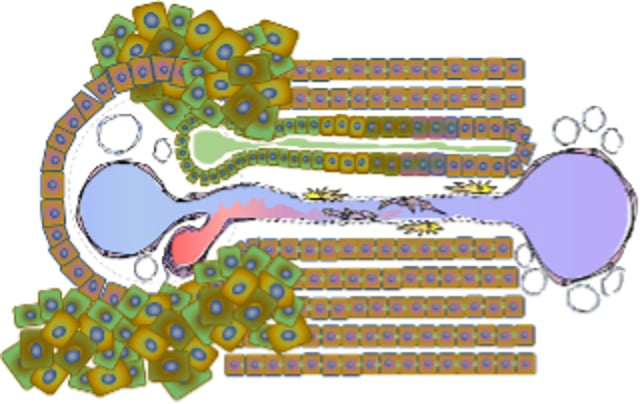
Portal tract with a ductular reaction where cholangiocytes (proliferative multicolored cells) without ductular structure disrupt the limiting plate and extend into adjacent hepatic parenchyma.
Courtesy of Dr. Sharon Center.
Disorders involving direct targeting of cholangiocytes or bile ducts are collectively referred to as cholangiopathies. Activated or reactive cholangiocytes elaborate proinflammatory and profibrotic chemokines and cytokines and frequently undergo remarkable cell proliferation, characterized as a ductular reaction. This phenomenon reflects rapid transformation of cholangiocytes into a proliferative-reparative population that can serve as either biliary or hepatocyte progenitors. Ductular reactions are a common histologic response pattern exhibited in diverse number of hepatobiliary disorders.

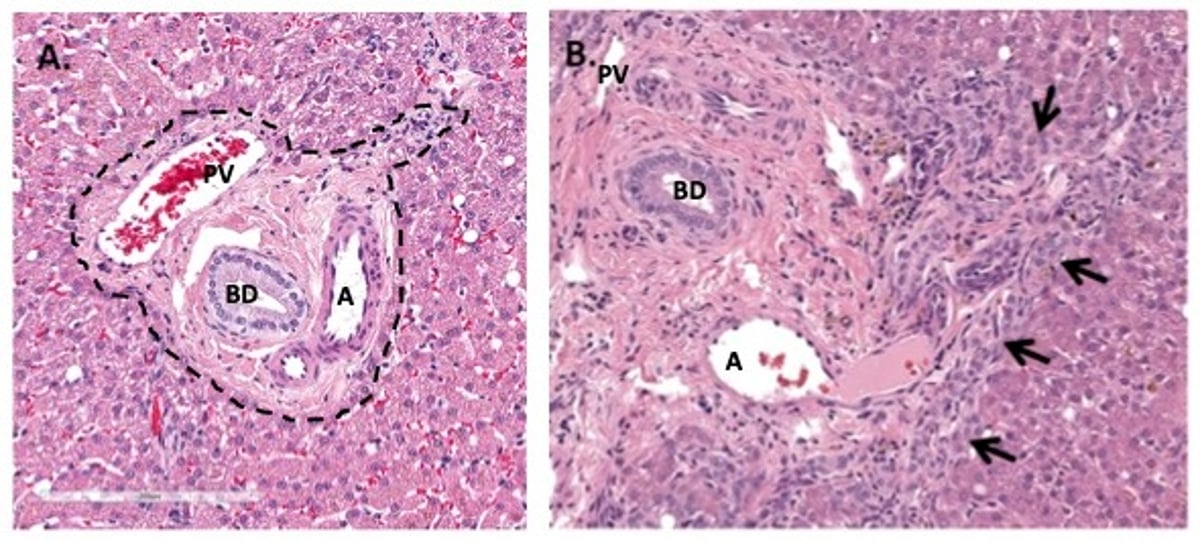
Photomicrographs of liver sections illustrating a normal portal tract (A) and a portal tract from a dog with a postnecrotic healing response (B) with a considerable ductular reaction (arrows). A—The limiting plate is marked with a dashed line. B—The limiting plate is disrupted and cannot be ascertained due to the considerable ductular reaction. PV = portal vein. A = artery. BD = bile duct. H&E stain.
Courtesy of Dr. Sharon Center.
This response is characterized by a disorganized proliferative population of cholangiocytes at the periphery of portal tract margins (limiting plate region), lacking recognizable ductal morphology. When extreme, reactive cholangiocytes breach the portal tract extending into the lobule, exhibiting a portal-to-portal bridging orientation. Ductular reactions can complicate histologic interpretation of liver biopsies because they generate confusion about the involvement of duct-targeting inflammatory response (ie, cholangitis).
Diseases disrupting cholangiocyte structure or function or canalicular bile transport lead to bile flow stasis (cholestasis). Cholestasis can cause accumulation of canalicular bile, distinguished microscopically by the accumulation of yellow-tan-green globular material at the intercellular junction between two hepatocytes.
Canalicular bile casts or plugs are commonly associated with gram-negative bacterial infections because endotoxin disrupts bile pump transporter expression.
Canalicular bile casts develop in a diverse number of hepatobiliary disorders and are especially notable on cytologic inspection of liver aspirates. Thus, cytologic identification of bile casts in liver aspirates has limited value for prognosis or definitive diagnosis.
Canalicular cholestasis and jaundice caused by endotoxemia or bacterial infection may be exhibited in the absence of increased liver enzyme activity. Canalicular bile casts reflecting a primary hepatobiliary disease usually are associated with increased liver enzyme activities (particularly associated with cholestatic enzymes ALP and GGT) and histologic injury to the biliary tree or severe canalicular cholestasis on routine H&E staining.
Disorders directly involving bile ducts are collectively categorized as cholangiopathies (eg, cholangitis denoting intrahepatic bile duct inflammation, cholangiohepatitis denoting inflammation affecting intrahepatic bile ducts and adjacent hepatic parenchyma, as well as disorders imposing mechanical bile duct obstruction). Inflammation of the largest bile ducts external to the liver (eg, extrahepatic large hepatic ducts or common bile duct) is denoted as choledochitis.
In disorders compromising the integrity of the canalicular barrier or that redesignate bile transporter expression, bile may spill into the ultrafiltrate, sinusoidal blood, or portal regions, directly contaminating systemic blood and causing jaundice. Direct cell membrane exposure to bile is injurious, with the exception of normal exposure of the intraluminal biliary epithelium.
Bile formation is a unique and essential liver function. Bile represents a complex aqueous secretion, originating in hepatocytes, that is modified during transit in the biliary tree by absorptive and secretory processes. Bile production and transport run countercurrent to sinusoidal circulation. Bile is either transported to the gallbladder where it is concentrated for efficient mealtime delivery to the intestinal lumen, or bile bypasses the gallbladder and is directly excreted into the duodenum.
Initially, canalicular bile has a high water content (~95%) and contains a multitude of endogenous constituents (including bile salts [bile acids], bilirubin, phospholipid, cholesterol, amino acids, steroids, enzymes, porphyrins [heme-associated pigments], vitamins, and heavy metals) as well as exogenous constituents including drugs, xenobiotics and toxins.
Many functions have been attributed to bile:
functioning as an indispensable excretory route for potentially harmful exogenous lipophilic substances and certain endogenous substrates (ie, bilirubin and bile salts) with large molecular weights (> 300–500 daltons) precluding renal filtration and not promptly eliminated by renal tubular excretion
distribution of bile salts, the major organic solute in bile that is physiologically indispensable for emulsification and assimilation of dietary fat and fat-soluble vitamins
functioning as the principal route of cholesterol elimination
providing essential intestinal distribution of biliary IgA that protects against enteric pathogen translocation
conservation of enteric commensal over opportunists and pathogenic bacteria, providing some degree of protection against enteric dysbiosis
distribution and elimination of inflammatory cytokines and chemokines, which can be either protective or injurious
stimulating the innate immune response and upregulating defensive cytokines
enabling cholehepatic and enterohepatic substrate circulation, the prime example being bile acids, now classified as hormones because of their complex interactions with the bile acid receptor—a nuclear receptor, also known as the farnesoid X receptor (FXR)—and metabolism
initiating gallbladder relaxation during the interdigestive interval, enabling its function as a low-pressure bile reservoir (via FXR and FGF19 signaling)
aiding in the removal or recovery of certain hormones and vitamins
Bile initially produced by canalicular membranes has a light yellow to green color. Major bile components include: conjugated bile acids, conjugated bilirubin, electrolytes, bicarbonate, mucin, phospholipids, cholesterol, immunoglobulin A (IgA), and molecules and substrates eliminated in bile, with some undergoing enterohepatic circulation.
Enterohepatic circulation denotes substances eliminated in bile that undergo enteric absorption and recirculation to the liver via the portal vein. The classic example of enterohepatic circulation is that associated with bile acid homeostasis.
Hepatic bile formation is categorized as bile acid dependent or independent. Primary bile acids are predominantly involved with bile acid–dependent bile flow. Bile acid–independent bile flow reflects transport of organic substrates, primarily glutathione (GSH) at the canalicular membrane and bicarbonate secretion by cholangiocytes.
Canalicular bile formation is a continuous process driven by transporters excreting organic solutes (eg, GSH, bile acids) that are passively followed by water and electrolytes and other organic substrates, such as glucose and amino acids. Concurrently, the canalicular membrane forms and extrudes lipid vesicles destined to form mixed micelles (aggregates of phospholipid, phosphatidylcholine [also called lecithin], free cholesterol, and bile acids). Lipid vesicles and mixed micelles attenuate the osmotic impact of and epithelial cytotoxicity of bile salts.
Transport of mixed micelles to the intestines is essential as these facilitate fat digestion and assimilation. High biliary cholesterol concentrations impose risk for cholesterol cholelithiasis (gallstones), a scenario extensively studied in humans in whom high prevalence of cholesterol gallstones exists. Fortunately, dogs and cats do not exhibit similar cholesterol bile saturation and do not share this morbidity.
This physiologic difference between humans and dogs and cats should be considered when contemplating use ursodeoxycholic acid for cholelith dissolution. The cholelith dissolution protocol, well documented in humans with cholesterol cholelithiasis, does not achieve similar response in dogs or cats because of differences in cholelith composition.
Bile acids are amphipathic organic anions (amphipathic = having a hydrophilic and lipophilic, detergent-like molecular interface) synthesized in the liver from cholesterol and now recognized to modulate numerous signaling cascades in the liver and intestines. Historically, they were of interest simply because they mediate the digestion and absorption of dietary fat and fat-soluble vitamins. Within the last decade, it has been clarified that bile acids function as vital regulatory ligands of bile acid synthesis and homeostasis, bile acid enterohepatic circulation, and diverse metabolic interactions of intermediary metabolism, as well as in adjusting the integrity of the intestinal barrier (ie, control of bacterial translocation).
Primary bile acids include cholic acid (cholate) and chenodeoxycholic acid (chenodeoxycholate). Secondary bile acids are derived in the gut from bacterial dehydroxylation of primary bile acids, with cholic acid yielding deoxycholic acid and chenodeoxycholic acid yielding lithocholic acid (a profoundly noxious and injurious bile acid). Most biliary bile acids exist in conjugated form and are incorporated into mixed micelles.
Bile acids are obligatorily conjugated with taurine in cats and either taurine or glycine in the dog. The conjugation process converts bile salts into stronger acids, limiting their passive reabsorption during biliary transport. Once entering the intestines in bile, bile acids undergo enterohepatic circulation, with efficient uptake first by enterocytes in the ileum and then by hepatocytes at their sinusoidal interface. Passive absorption of bile acids also occurs along the entire small intestine and colon but is less efficient than active uptake ileum.
High-efficiency transporters conserve bile acids such that there is a < 5% loss in feces per day. The bile acid pool recirculates ~4–12 times per day depending on the frequency of meal ingestion and the level of dietary fat ingestion. The small percentage of bile acids escaping active or passive absorption in the intestines is the fraction subject to bacterial modification; deconjugation, and dihydroxylation to secondary bile acids. These bile acids are either passively absorbed or eliminated in feces.
The tight regulation of the bile acids’ enterohepatic circulation is exploited in their use in diagnostic tests to detect cholestatic liver injury, severe parenchymal liver injury, advanced-stage liver disease, and hepatofugal portal circulation (perfusion “around” or “away” from the liver). Bile acids are also used to diagnosis microvascular dysplasia, a syndrome of congenital portal venous hypoperfusion in small-breed dogs. Bile acids in the circulating blood more sensitively reflect cholestatic liver injury than bilirubin and are extensively used to identify portosystemic shunting of blood around the liver (eg, congenital portosystemic shunts, acquired portosystemic shunts, congenital portal venous hypoperfusion associated with microvascular dysplasia in small-breed dogs).
Measuring bile acids in a hyperbilirubinemic patient with strong evidence of hepatobiliary disease would be superfluous. Occasionally, they are used to help differentiate hemolytic from hepatobiliary jaundice; however, usually that is obvious based on assessment of hematocrit.
Adjustment of the enterohepatic circulation and synthesis of bile acids is under the influence of the bile acid receptor, also known as the farnesoid X receptor (FXR) or NR1H4—a member of the nuclear receptor family (NHR; receptors exert transcription factor activity upon modulation by ligands). FXR is the nuclear receptor for bile acids and transcriptionally regulates several genes, adjusting bile acid homeostasis through four negative feedback circuits:
Hepatocyte FXR induces expression of transcription factor small heterodimer partner (SHP), NROB2, which represses other transcription factors. The SHP downregulates expression of the rate-limiting enzyme regulating bile acid synthesis, cholesterol 7 alpha-hydroxylase (CYP7A1).
In the ileum enterocytes, FXR induces expression of fibroblast growth factor 19 (FGF19), which is released into the portal vein. In the liver, this FGF19 interacts with the FGF receptor-4, also repressing CYP7A1 expression, further limiting bile acid synthesis.
In hepatocytes, FXR also adjusts two bile salt membrane pumps, resulting in a net decrease in hepatocyte bile acid concentration (FXR increases expression of the bile salt export pump [BSEP] that transports bile acids into the canaliculus; FXR suppresses expression of the sodium taurocholate co-transporting polypeptide [NTCP] that pumps bile acids into the hepatocyte).
In enterocytes of the ileum, FXR also downregulates the apical sodium-dependent bile acid transporter (ASBT) that plays a key role in the enterohepatic circulation of bile acids (this transporter moves bile acids from the gut lumen into the enterocyte) while concurrently upregulating an organic solute and steroid transporter (OSTa-OSTb), whose function it is to pump bile acids out of the enterocyte into the portal circulation. The net effect of these changes is to decrease the concentration of bile acids in these enterocytes.
Bile acid–dependent bile flow has a powerful influence at the canaliculus with non–micelle-forming bile salts imparting the greatest influence (eg, dehydrocholate). Once transported across the canalicular membrane, bile acids are promptly integrated into mixed micelles, achieving a marked concentration gradient (high to low) between bile and blood; bile acid concentrations in bile exceed plasma bile acid concentrations by 100- to 1,000-fold.
This extreme concentration gradient is why chemical peritonitis rapidly evolves when bile spills into the abdominal cavity. Formation of micelles allows cholesterol to remain in solution and lowers the free bile salt concentrations, greatly attenuating the toxic detergent effect of bile salts on biliary epithelium. It is this same detergent effect that is desirable for facilitation of lipid digestion and absorption.
Bile acid–independent bile flow largely involves GSH transport across canalicular membranes. The strong osmotic influence of GSH derives from its hydrophilic nature, active exportation across the canalicular membrane, and subsequent hydrolysis by membrane-affiliated gamma glutamyl transferase (GGT). Breakdown of GSH into its three constituent amino acids (cysteine, glutamate, glycine) triples its osmolar impact and drags water and electrolytes with it into bile (via paracellular pathways). This is why thiol donors (substances providing thiol group-bearing cysteine) are used as choleretics (drugs that stimulate bile flow)—particularly relevant in animals with cholestatic liver disease at risk for low hepatic GSH concentrations.
Because GSH is not directly transported across cell membranes, oral GSH products function as an expensive way to supplement cysteine, the GSH constituent amino acid (bearing -SH groups) that is rate limiting to GSH synthesis. Rather, administration of S-adenosylmethionine (SAMe, using bioavailable forms) has more logical benefit because SAMe functions as a cysteine donor (and thus, a GSH donor) and also supports methylation reactions essential to intermediary metabolism, along with additional benefits.
Lack of sufficient SAMe synthesis contributes to the low hepatic GSH status of patients with liver disease (proven in humans and experimental studies in other species, but yet to be definitively shown in dogs or cats).
A choleretic influence of SAMe was demonstrated in gallbladder bile in clinically normal cats treated with high-dose bioavailable SAMe (40–60 mg/kg, PO, q 24 h).
For acute in-hospital GSH support for animals with critical hepatobiliary disease (ie, fulminant hepatic failure, advanced-stage liver disease, awaiting resolution of EHBDO secondary to pancreatitis, or preparation for biliary decompression surgery), IV-administered N-acetylcysteine is used as a thiol donor. This should be transitioned to SAMe when oral medications are possible.
Canalicular bile is modified by biliary epithelium during transit through the intrahepatic and extrahepatic biliary tree and during storage in the gallbladder. During transport through intrahepatic biliary elements, is modified by secretion and reabsorption of fluid and inorganic electrolytes. Cholangiocytes also uptake bile salts (Na1-dependent transporter) as an initial step in a cholehepatic shunt pathway involved with the intrahepatic recycling of bile salts through a periductular capillary plexus. Under normal conditions, high intraductular bile salt concentrations (millimolar range) negate the impact of this process.
Cholehepatic shunting has physiologic importance in signaling ductular mucin production and bicarbonate secretion and is responsible for the extreme serum-bile acid concentrations exhibited in many cholestatic disorders. Bile ducts substantially contribute to bile formation and flow through bicarbonate production, stimulated by secretin (a hormone derived from duodenal and jejunal mucosa). Ductal secretion contributes to between 10% and 40% of basal bile flow, depending on the species. In humans, 40% of bile volume has a ductular origin.
The yellow-green color of bile reflects eliminated bilirubin pigments. Disorders causing cholestasis, panlobular liver injury and dysfunction, or excessive heme release, exceeding the limits of bilirubin clearance, can lead to development of jaundice.





