



Foot-and-mouth disease is one of the world's most economically important viral diseases of livestock. The virus infects cattle, pigs, sheep, goats, and many cloven-hoofed wildlife species. The infection results in vesicular lesions in and around the mouth and on the feet, and animals become reluctant to eat or move. Confirmation of the infection requires laboratory analysis, usually involving real-time RT-PCR assay, and serotyping is achieved using antigen ELISA or nucleotide sequencing. Inactivated virus vaccines are available.
Foot-and-mouth disease (FMD) is a highly transmissible disease caused by infection with an Aphthovirus, a member of the family Picornaviridae. There are 7 serotypes of the virus, termed: O, A, C, Asia 1, and SAT 1, SAT 2, and SAT 3 (southern African territories serotypes 1, 2, and 3). Serotype C may be extinct in the wild. Further diversity is found between strains within each serotype.
The virus primarily affects cloven-hoofed animals of the order Artiodactyla. Livestock hosts include cattle, pigs, sheep, and goats. FMD virus has also been reported to affect many wild artiodactyls, including African buffalo, bison, giraffes, Bactrian camels, deer, and antelope. Asian elephants (order Proboscidea) are also susceptible.
FMD is characterized by fever and vesicles in the mouth and on the muzzle or snout, teats, and feet of animals and is transmitted most easily through direct contact with infected animals or their excretions. The virus can also be transmitted as an aerosol via respiratory secretions and through milk, semen, and ingesting feed that contains meat from infected animals or that has been contaminated with their secretions.
In a susceptible population, morbidity can reach 100%; however, fatalities are rare except in young animals or when lesions become secondarily infected by bacteria.
FMD is distributed worldwide but has been eradicated from North and Central America, most of South America, Europe, and Australia. It has not been reported in New Zealand and Antarctica.
In endemic regions (including most of Africa, the Middle East, and southern and southeastern Asia), FMD has a substantial impact on agricultural productivity and places economic constraints on international trade of livestock and animal products (see figure showing ).


Global distribution of foot-and-mouth disease. Distinct pools of viruses, including various serotypes, circulate in different regions of the world, as indicated. (2026)
Courtesy of Dr. Antonello Di Nardo, Vesicular Disease Reference Laboratory, The Pirbright Institute.
The disease can be easily reintroduced into FMD-free areas unless strict precautions are in place. Outbreaks in FMD-free areas can be severely disruptive and require substantial resources to control, as in the 2001 epizootic in the UK. This outbreak, which lasted for approximately 8 months, is estimated to have cost approximately $10 billion (USD). Major components of the cost of outbreaks in free areas include lost trade and the large numbers of animals culled to limit spread of the disease.
Smaller outbreaks occurred in Germany, Hungary, and Slovakia in 2025. They were quickly contained; however, trade losses were nevertheless substantial.
Epidemiology and Transmission of Foot-and-Mouth Disease
The different serotypes of FMD virus (FMDV) are not uniformly distributed within the endemic regions. Serotype O is responsible for the majority of outbreaks globally.
Six of the 7 serotypes have occurred in Africa (O, A, C, SAT1, SAT2, SAT3), 6 in Asia (O, A, C, Asia 1, SAT 1, and SAT 2), and 3 in South America (O, A, C). Most of the Americas, Australia, New Zealand, and Europe are now normally free of FMD (the last outbreaks in Europe were in Germany, Hungary, Slovakia, and Cyprus in 2025).
No infection caused by serotype C anywhere in the world has been reported since 2004, and it might now be eradicated outside of the laboratory. However, vaccination against this serotype still occurs in a few places, so the complete absence of the virus in the field is difficult to prove.
FMD virus is transmitted via various routes:
direct contact with infected animals
indirect contact with secretions or excretions (including semen and milk) from infected animals
mechanical vectors (people, horses, dogs and cats, rodents, birds, vehicles, and equipment)
air movement over land and water
The virus can enter the host via inhalation, ingestion, or skin wounds and mucous membranes. Breeding is a possible route of transmission for the SAT viruses in African buffalo populations.
A potential scenario for introduction of the virus into a previously FMD-free area is for a susceptible population, such as pigs, to be given imported food derived from an infected animal. Virus then spreads from pigs, which can exhale a thousandfold more virus than cattle, to more susceptible cattle hosts via aerosol. Virus was reported to have traveled over water > 250 km (> 150 miles) from Brittany, France, to the Isle of Wight, UK, in 1981; however, it usually travels no more than a few kilometers over land (1, 2).
FMD has high agroterrorism potential because of its infectivity, stability in the environment, and potential to cause enormous economic losses and societal disruption.
Humans can act as mechanical vectors of FMD by carrying virus on clothing or skin; however, they can greatly decrease the risk of inadvertent transmission by showering and changing clothes.
FMD is not a zoonosis and is not considered a public health problem.
FMD virus is environmentally resistant but can be easily inactivated outside the pH range 6–9, by desiccation, and at temperatures > 56°C (133°F). It is resistant to lipid solvents such as ether and chloroform; however, sodium hydroxide (lye), sodium carbonate (soda ash), citric acid, and acetic acid (vinegar) are effective disinfectants. Iodophors, quaternary ammonium compounds, hypochlorite, and phenols are less effective, especially in the presence of organic matter (3).
FMD virus is shed into the milk of dairy cows before clinical signs develop. Infectivity is substantially decreased by heat treatment of milk; however, trace amounts of FMDV could survive, depending on the method (4).
FMD virus can survive on hay or straw bedding for up to 20 weeks, in dry fecal matter for up to 14 days in summer, in fecal slurry for up to 6 months in winter, and in soil for 3 (summer) to 28 (winter) days. However, the extent of virus survival in these materials is dependent on the initial level of contamination (5).
Properties of Foot-and-Mouth Disease Virus
Foot-and-mouth disease virus, like other picornaviruses, has a positive-sense RNA genome. The viral genome is approximately 8,500 nucleotides in length and includes a single large open reading frame that encodes a large polyprotein (approximately 2,300 amino acids in length). During and after the synthesis of this polyprotein, it is processed, largely by virus-encoded proteases, to generate 15 mature proteins.
The structural proteins of the virus (termed VP1, VP2, VP3, and VP4) are produced from the capsid precursor P1-2A (see ). The near-spherical virus particle (approximately 25–30 nm in diameter) includes 60 copies of each of the structural proteins, plus a single copy of the virus genome. The proteins VP1, VP2, and VP3 are exposed on the outside surface of the virus, whereas VP4 is entirely internal.

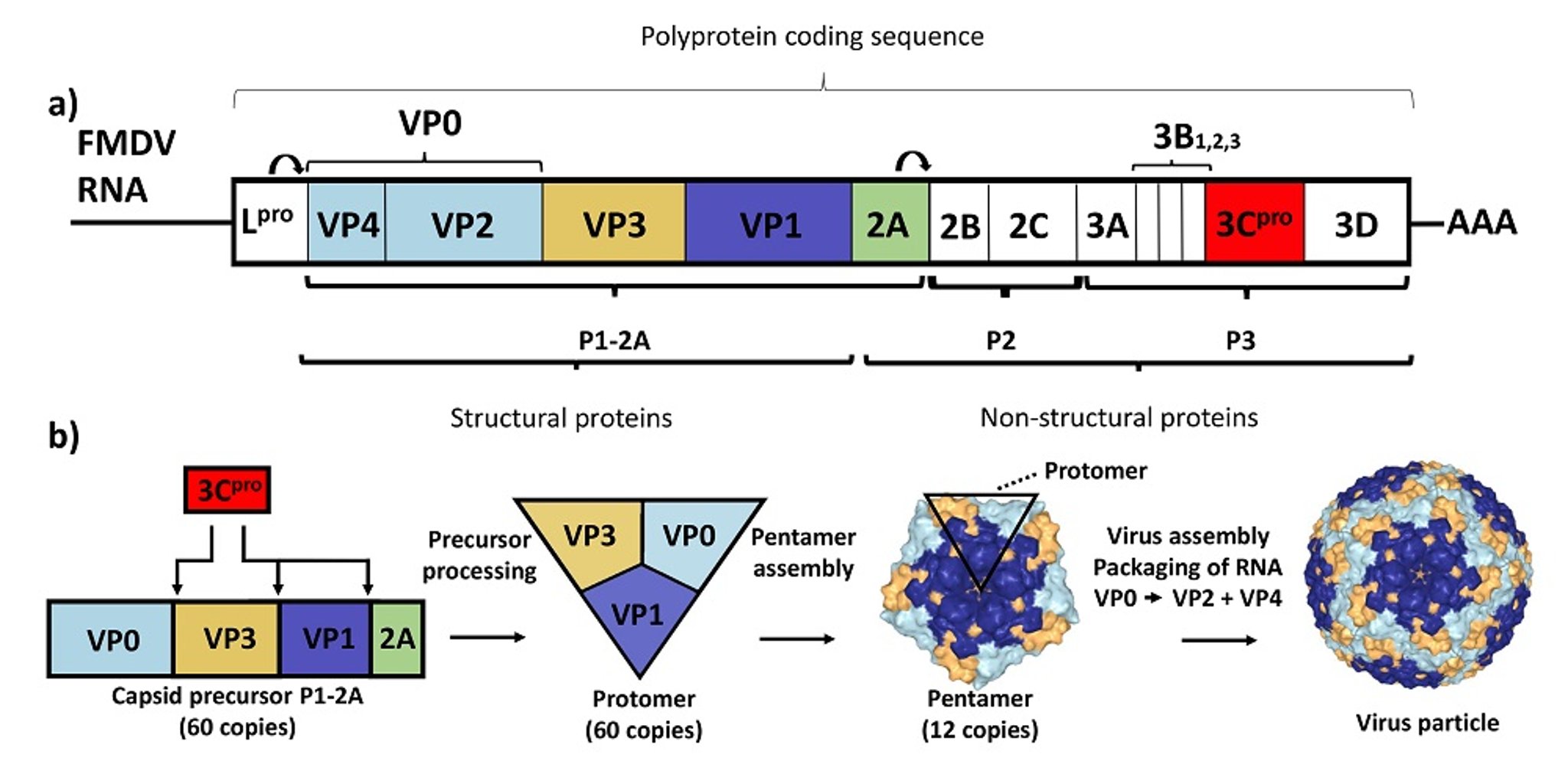
a) The foot-and-mouth disease virus RNA genome contains a single open reading frame encoding a polyprotein that is processed to yield 15 mature products and a variety of precursors. b) The structural proteins, which make up the virus particle, are produced by processing the capsid precursor (P1-2A), and the products (60 copies of each) assemble with a single copy of the RNA genome into new virus particles.
Courtesy of National Veterinary Institute, DTU Vet, Lindholm, Denmark.
The virus capsid serves to protect the RNA genome while it is outside of a host cell and also facilitates entry into cells by binding to specific receptors on the cell surface.
After virus internalization, the RNA genome is released into the cytoplasm of the cell. It is translated directly to make the viral proteins, including various nonstructural proteins, and then the RNA genome is replicated (via a negative-sense intermediate) by some of these nonstructural proteins.
Packaging of the positive-sense RNA by the capsid proteins generates new virus particles. Many thousands of new virus particles can be produced in an infected cell within a few hours.
Pathogenesis of Foot-and-Mouth Disease
The primary sites of infection and replication of foot-and-mouth disease virus are the mucosa of the nasopharynx in cattle and the crypt epithelium of the oropharyngeal tonsils in pigs. The virus can also enter through skin lesions.
Once distributed by the lymphatic system and blood, the virus replicates in the epithelium of the mouth, muzzle or snout, teats, feet, and areas of damaged skin (eg, the elbows and hocks of pigs). Vesicles develop quickly and rupture within 48 hours.
More than 50% of ruminants that recover from FMD (including those that were vaccinated and later exposed to the virus) can become carriers—ie, they maintain low levels of infectious virus in their pharyngeal region long after the acute phase of infection. The carrier state can last for a few years in cattle, several months in sheep, and > 5 years in African buffalo (6). Strikingly, such persistent infections do not occur in pigs.
The risk posed by carrier animals seems low (but may not be zero) because it has not been possible to transmit FMD disease (under controlled conditions) from carrier cattle to naive cattle or other susceptible animals by close contact for extended periods of time. However, transmission of the disease has been achieved from carrier African buffalo to cattle and also by direct artificial transfer of invasively collected pharyngeal fluid from carrier cattle to naive cattle.
The incubation period of FMD is variable and depends on the host, environment, route of exposure, and virus strain. After infection with FMD virus, the typical incubation period in sheep and goats is 3–8 days, ≥ 2 days in pigs, and 2–14 days in cattle. The incubation period can be as short as 18 hours for host-adapted strains in pigs, especially under intense direct contact (7).
Animals can transmit the virus before the appearance of clinical signs because the virus replicates in the pharynx or mouth before disease or even viremia is observed.
Clinical Findings of Foot-and-Mouth Disease
Clinical signs of foot-and-mouth disease in cattle include fever of approximately 40°C (104°F), followed by vesicular lesions appearing on the tongue, hard palate, dental pad, lips, gums, muzzle, coronary band, interdigital cleft, and teats in lactating animals. Acutely affected individuals can salivate profusely and make characteristic smacking noises, stamp their feet, and prefer to lie down. Ruptured oral vesicles can coalesce and form erosions but heal rapidly less than two weeks after vesicle formation.
See images of clinical signs in cows (; ; ; ; ; and ) and in pigs (; ; foot lesion, and ).


An experimentally inoculated cow on day 3 after infection shows profuse salivation and nasal discharge, acute clinical signs of the foot-and-mouth disease complex. At this stage, virus and viral RNA are readily detectable in an oral swab or saliva sample.
Courtesy of National Veterinary Institute, DTU Vet, Lindholm, Denmark.

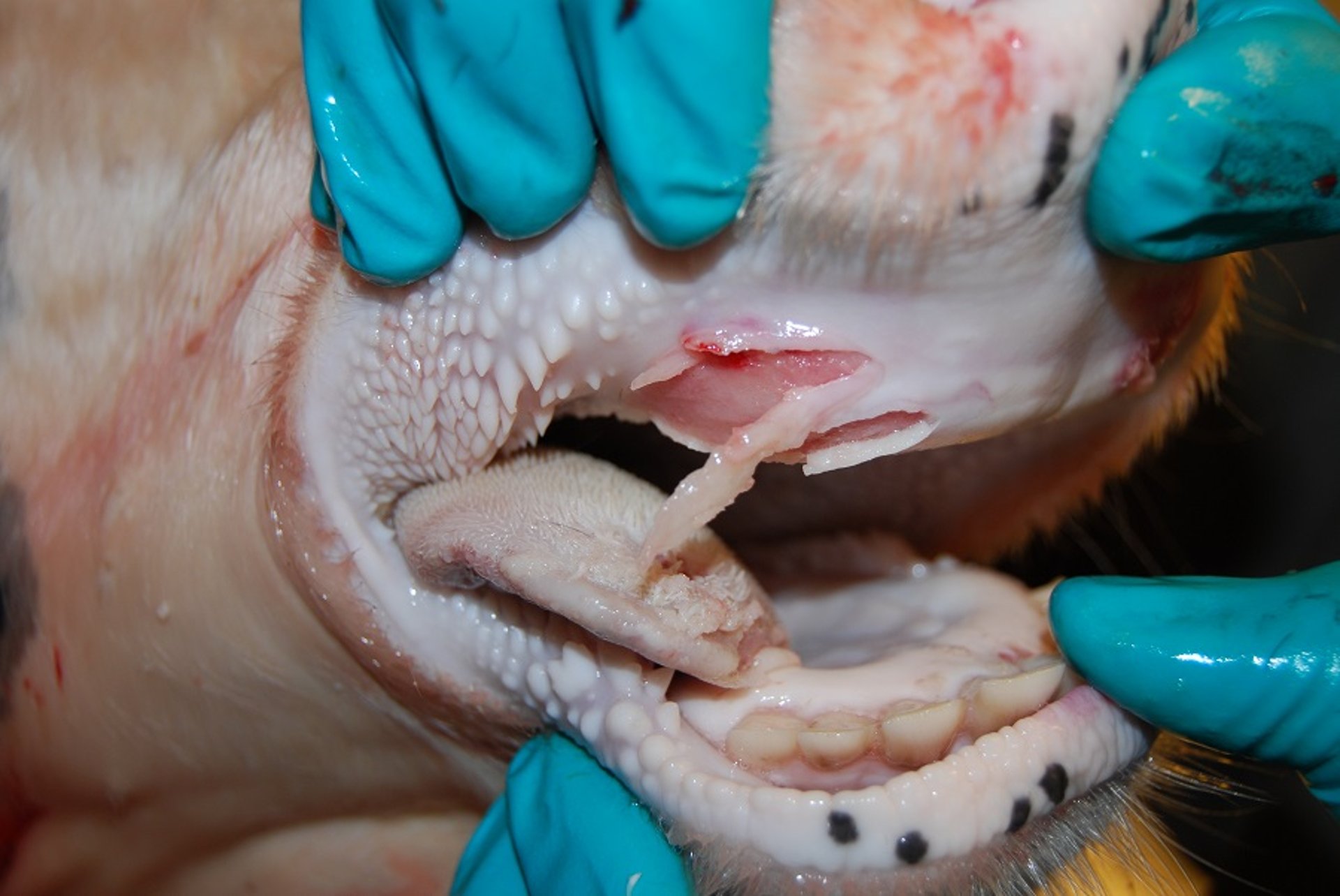
Erosive lesions of foot-and-mouth disease on the dental pad and tongue of a cow. Vesicles have ruptured, and the overlying epidermis has sloughed off. Hyperemia and hemorrhage in the underlying tissue follows. This is a contact animal on day 10 after experimental exposure. The sharp demarcation of the edges of the vesicle on the dental pad is indicative of recent rupture. This is a fresh lesion highly suitable for sample collection.
Courtesy of National Veterinary Institute, DTU Vet, Lindholm, Denmark.

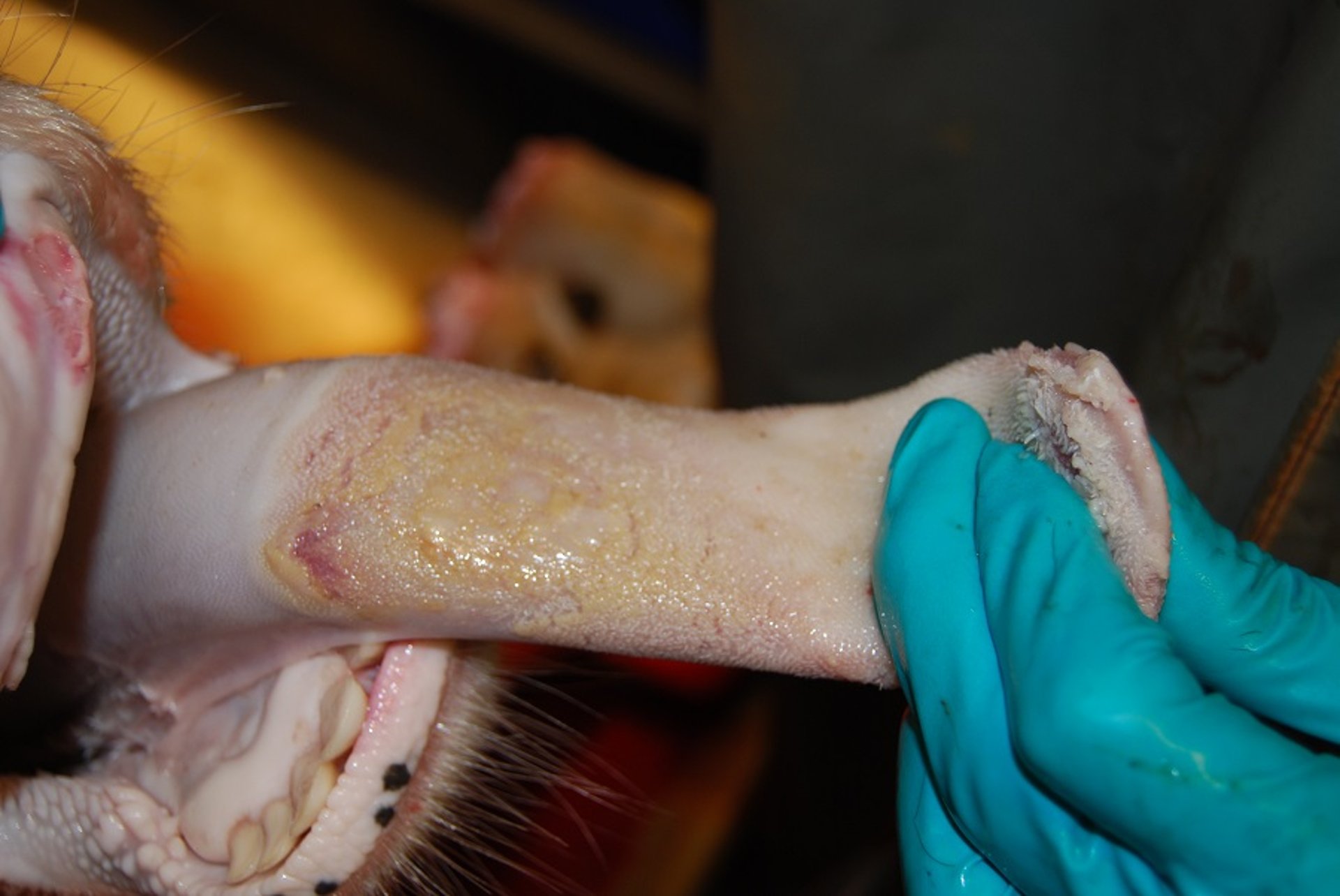
Healing foot-and-mouth disease lesion on the dental pad of a cow. This was a contact animal on day 10 after experimental exposure. Abundant fibrin deposition indicates an old lesion. Material collected from such lesions often contains only small amounts of viral RNA, and there might not be any infectious virus at all.
Courtesy of National Veterinary Institute, DTU Vet, Lindholm, Denmark.
Vesicles on the feet of animals with FMD take longer to heal and are susceptible to secondary bacterial infection leading to chronic lameness or, in severe cases, death. Lesions can involve one or more feet.
Secondary bacterial mastitis is common due to infected teat vesicles resulting in resistance to milking. After vesicular disease develops, cattle quickly lose condition and milk production diminishes, which can persist chronically.
Occasionally, young calves can die without prior clinical signs of illness because of virus-induced damage to the developing myocardium.
Healing foot-and-mouth disease lesion on bovine tongue. Contact animal on day 10 after experimental exposure. Abundant fibrin deposition indicates an old lesion. Material collected from such lesions often contains only small amounts of viral RNA, and there might be no infectious virus at all.
Courtesy of National Veterinary Institute, DTU Vet, Lindholm, Denmark.
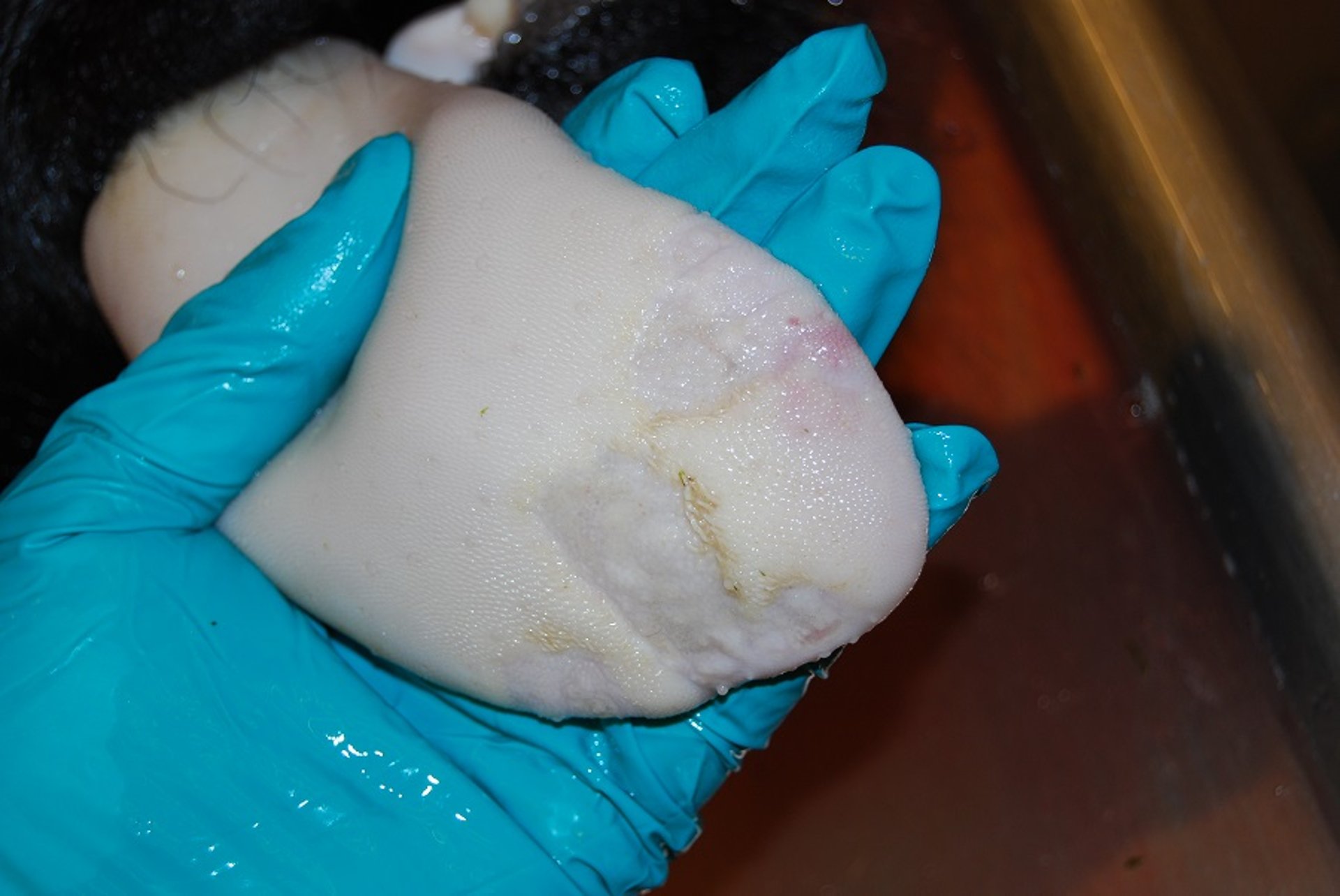
Almost fully healed foot-and-mouth disease lesion on bovine tongue. Contact animal on day 10 after experimental exposure. Reepithelialization is complete. It is unlikely that this lesion has any detectable viral RNA or infectious virus.
Courtesy of National Veterinary Institute, DTU Vet, Lindholm, Denmark.
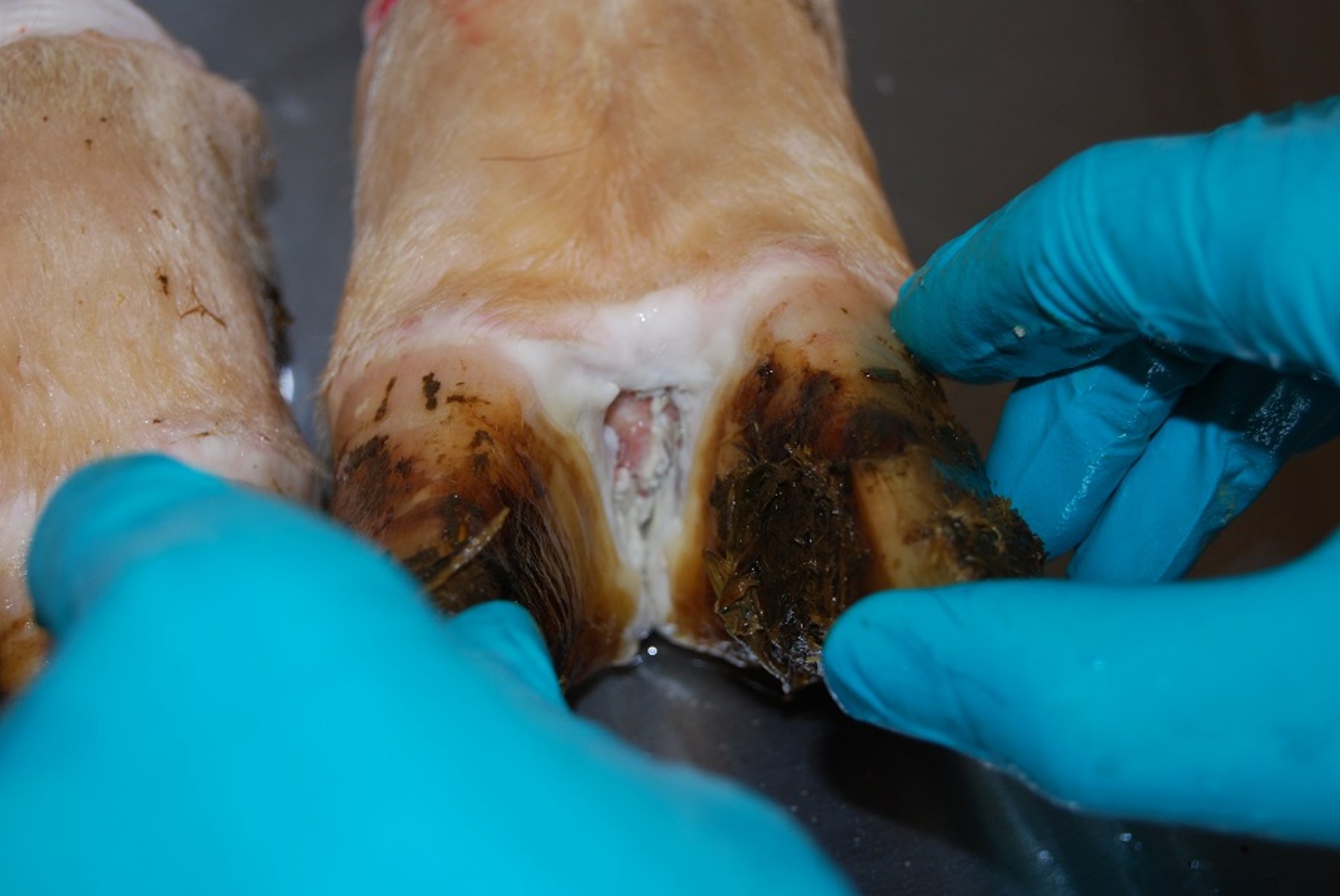
Erosive foot-and-mouth disease lesion in the interdigital cleft of a cow. This was a contact animal on day 4 after experimental exposure.
Courtesy of National Veterinary Institute, DTU Vet, Lindholm, Denmark.
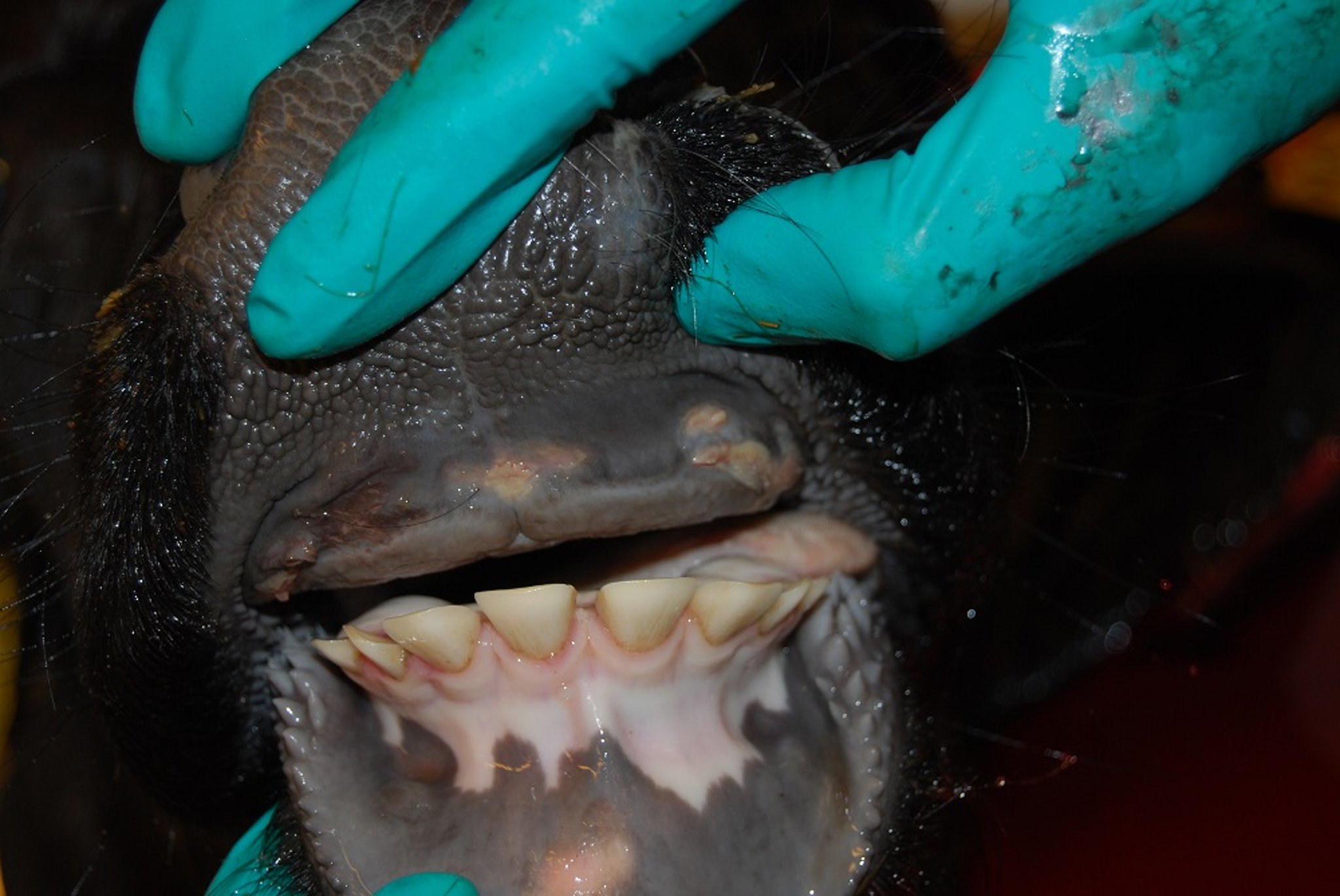
Pigs infected with FMD virus show mild to severe lameness and blanching around the coronary bands and can develop a fever of up to 41.5°C (107°F). Affected pigs become lethargic, huddle among other pigs, and may have little interest in feed. Vesicles develop on the coronary band and the heel of the foot (including accessory digits), the snout, mandible, and tongue. Additional vesicles can form on the elbows and hocks of pigs housed on rough surfaces. Depending on the severity of vesicles, the horn of the foot may completely slough off and cause chronic lameness in recovered pigs.
Young pigs, < 14 weeks old, may die without clinical signs of illness because of virus-induced myocarditis; this is more frequent in pigs than in calves.

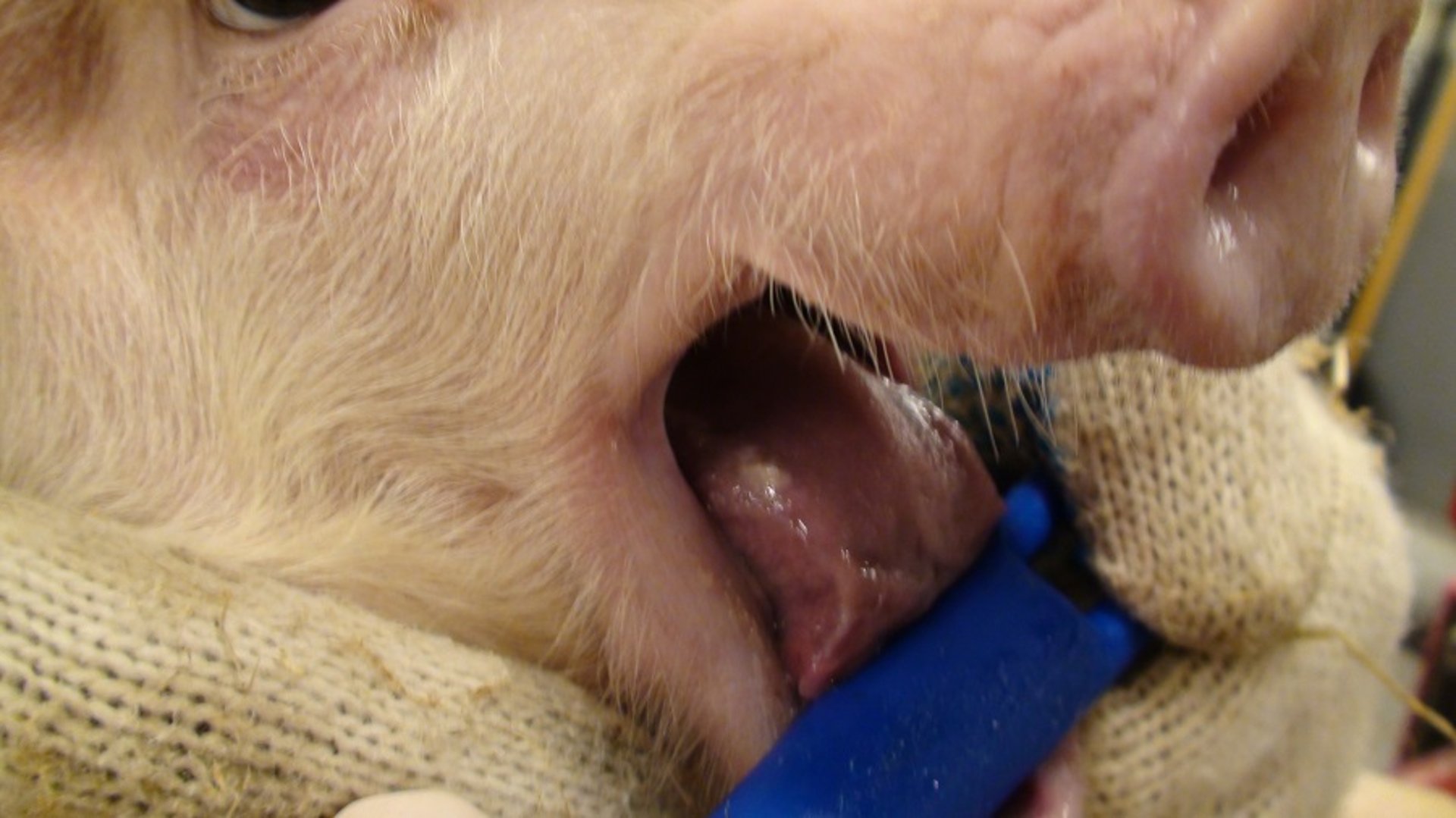
Unruptured vesicle on a pig's tongue. Experimentally inoculated animal on day 2 after infection with foot-and-mouth disease virus.
Courtesy of National Veterinary Institute, DTU Vet, Lindholm, Denmark.

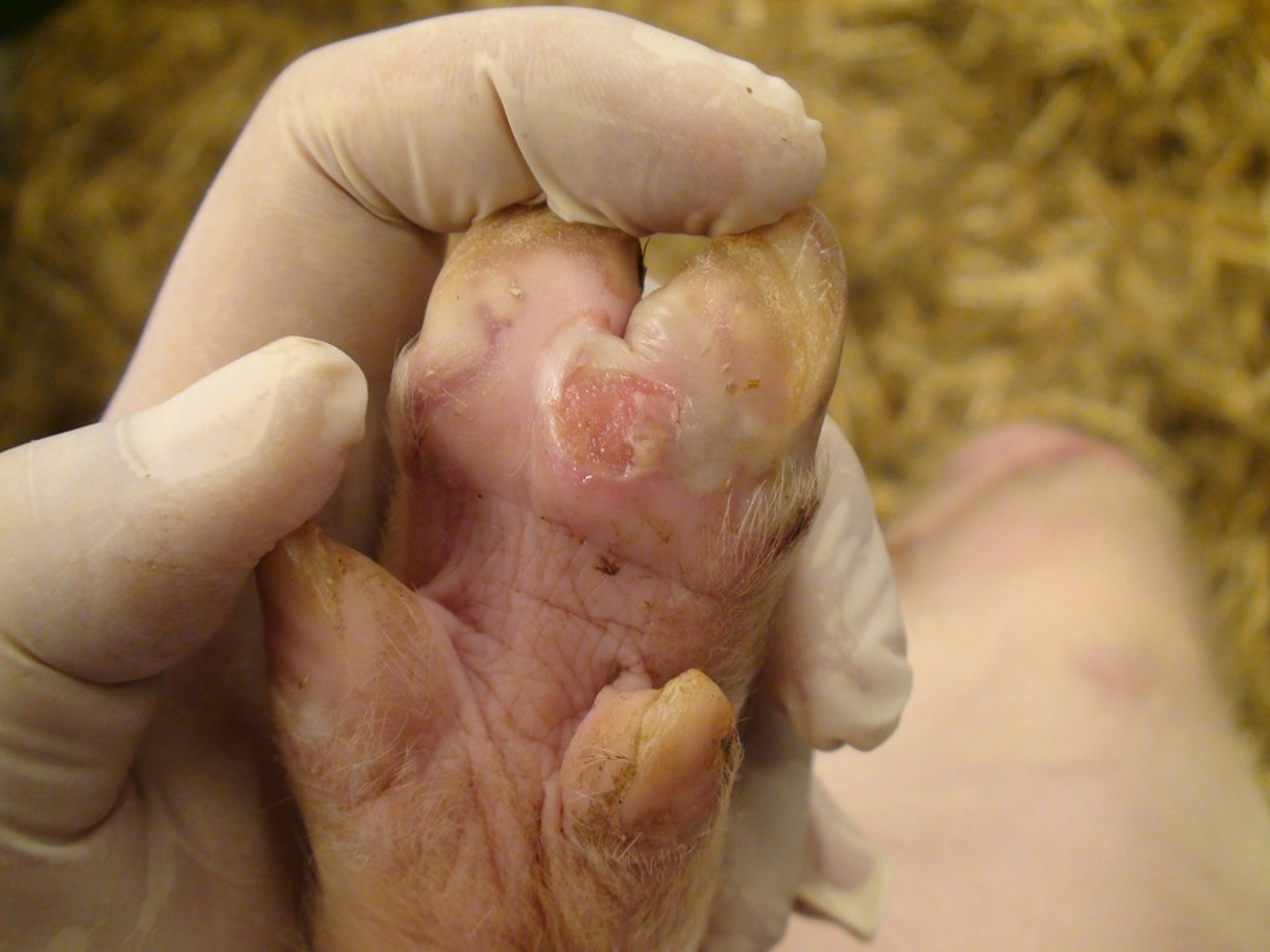
Foot lesions in an inoculated pig on day 4 after experimental infection with foot-and-mouth disease virus. Ruptured and white (unruptured) vesicles are present on the heel bulb. Lesions on the feet, resulting in lameness and reluctance to move, are a characteristic finding in pigs.
Courtesy of National Veterinary Institute, DTU Vet, Lindholm, Denmark.
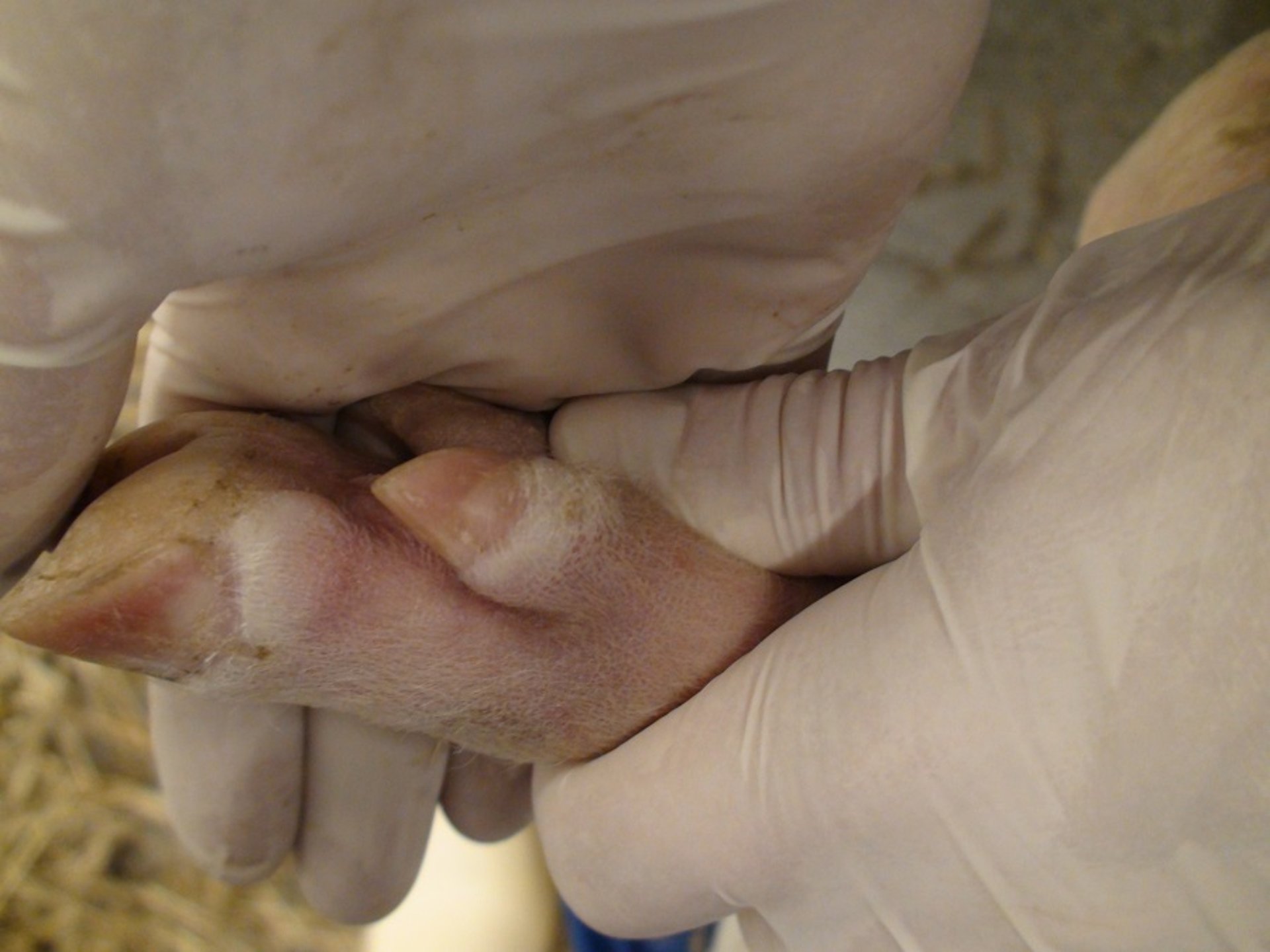
Foot lesion in a pig with foot-and-mouth disease. Note the blanching and swelling around the coronary band. Erosive damage of the skin will follow (see the same animal in the following photo). Contact animal on day 5 after experimental exposure.
Courtesy of National Veterinary Institute, DTU Vet, Lindholm, Denmark.
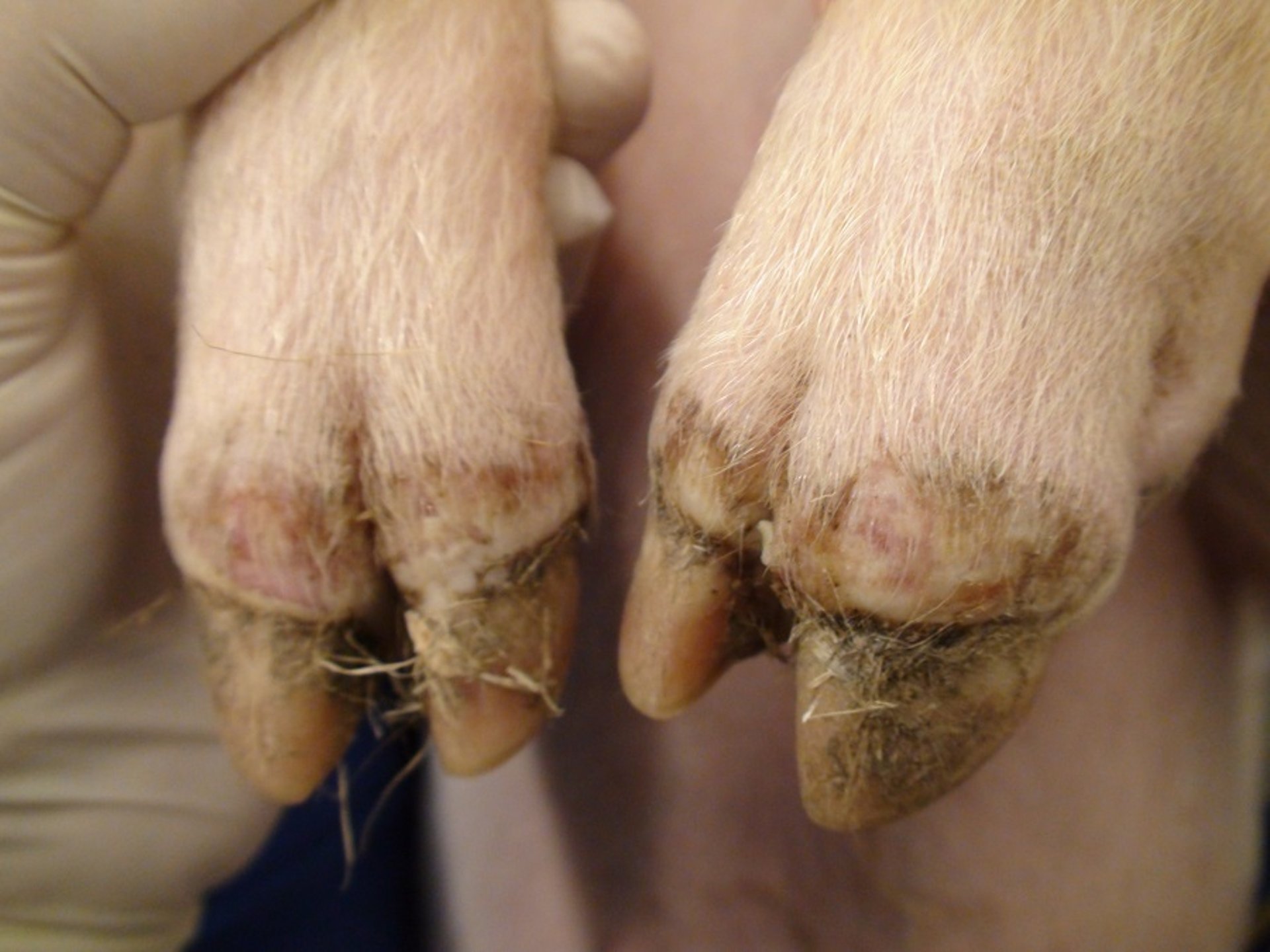
Foot lesions in a pig with foot-and-mouth disease. Note the erosive lesions on the coronary band (same animal as in previous photo). Secondary bacterial infection can occur. Contact animal on day 8 after exposure within an experiment.
Courtesy of National Veterinary Institute, DTU Vet, Lindholm, Denmark.
Clinical signs of FMD in sheep and goats can be largely inapparent. However, lameness is usually the first, if nonspecific, clinical sign of FMD infection observed in sheep and goats. This is followed by fever and vesicular development on the interdigital cleft, heel bulbs, coronary band, and mouth. Vesicles can also form on the teats of lactating animals and rarely on the vulva and prepuce.
Secondary infections result in decreased milk production, chronic lameness, and predisposition to other viral infections, including sheeppox and goatpox and peste des petits ruminants. Infection in immature sheep and goats, similar to that in young pigs, can cause heart failure and result in death without clinical signs.
Experimentally infected camelids are commonly reported to have mild, if any, clinical illness but can have severe infections resulting in salivation and mouth lesions and sloughing of the footpad and skin of the tarsal and carpal joints. Only Bactrian (two-humped) camels are susceptible to FMD; dromedary camels are not.
FMD infections in wildlife resemble clinical illness in their domestic counterparts; however, more severe lesions, such as sloughing of antlers or toe horn, are reported. Severe disease in Asian elephants has been reported, but African elephants appear not to be susceptible.
Aging of lesions is an important part of the epidemiological investigation of an FMD outbreak. Government agencies and professional societies have produced brochures that can help practitioners estimate the age of clinical lesions of FMD. These are freely available online from groups in both the US and Europe.
Diagnosis of Foot-and-Mouth Disease
Real-time RT-PCR assay, virus isolation, ELISA, or lateral flow devices for antigen detection
ELISA or virus neutralization tests for antibody detection
In cattle and pigs, clinical signs of FMD are indistinguishable from those of vesicular stomatitis, and in pigs they resemble clinical signs of swine vesicular disease, vesicular exanthema, and Seneca Valley virus disease. Other diseases that can be clinically similar include malignant catarrhal fever, mucosal disease, bluetongue, epizootic hemorrhagic disease, and contagious ecthyma (orf). Therefore, any vesicular disease in cloven-hoofed animals requires a diagnostic investigation by a suitable laboratory.
The tissue of choice for FMD virus isolation is vesicular epithelium from a fresh lesion. Vesicular fluid contains large amounts of virus as well; however, it is very difficult to collect in practice. At least 1 g of epithelium should be placed in a transport medium of phosphate-buffered saline solution or equal parts glycerol and phosphate buffer at pH 7.2–7.6. Samples should be kept refrigerated or transported on ice.
If vesicles are not present, oropharyngeal fluid can be collected via probang cup. During acute FMD infection, the virus and viral RNA are also readily detectable in saliva and on oral or nasal swabs.
Serum samples can also be tested (WOAH Terrestrial Animal Health Code 2024), but viremia is fairly short-lived (a few days); thus, when lesions are healing, FMD virus has already been cleared from the blood, and only antibodies can be detected. Repeated sampling of oropharyngeal fluid might be necessary to identify a carrier, because virus presence in such animals is low and fluctuates.
Primary laboratory diagnosis of FMD is usually performed by real-time RT-PCR assay; two separate assays targeting different regions of the RNA genome are commonly used. These assays are very sensitive and can detect FMDV genome even in poorly stored samples when virus infectivity has been lost. In epithelial homogenate prepared from fresh lesions, the presence of virus can also be demonstrated using an antigen ELISA, and this can determine the serotype.
In reference laboratories, sequencing of part of the genome (encoding the capsid proteins, particularly VP1) is frequently performed to determine the serotype and lineage of the strain. Concurrent virus isolation may be performed in appropriate cell culture systems. Commercially available lateral flow devices for rapid detection of virus antigen at the pen-side have proven useful in endemic settings. The devices' use in FMD-free areas must be carefully controlled to ensure that their lower sensitivity compared to real-time RT-PCR detection of viral RNA does not compromise the rapid detection of a virus intrusion. Negative results obtained with a lateral flow device always require laboratory confirmation with a more sensitive test. In FMD-free countries, lateral flow devices for FMD detection should be used exclusively by the veterinary services and not by private veterinarians or farmers.
Serological tests for FMD are used to certify animals for trade, to confirm suspected cases of FMD, test efficacy of vaccination, and provide evidence for absence of infection. Test cutoffs may be set at different levels for herd-based surveillance versus certifying freedom of infection for trade purposes.
The choice of serological test depends on the vaccination status of the animals.
Serological tests for antibodies against the structural (capsid) proteins of the virus (neutralization tests or ELISAs) cannot reliably detect infection in vaccinated animals because FMD vaccines induce antibodies against these proteins.
Pearls & Pitfalls
|
ELISA detection of antibodies against the nonstructural proteins, which are produced only during virus replication, can be used to determine past or present infection with any of the 7 serotypes, whether or not the animal has been vaccinated. However, ELISA detection of antibodies against the nonstructural proteins is less sensitive than serological tests for antibodies against the structural (capsid) proteins of the virus and can result in false negatives in cases with limited virus replication, such as vaccinated animals that become infected, because the vaccine suppresses viral replication (8).
Legacy methods such as agar gel immunodiffusion or complement fixation might still be used in resource-constrained settings or for special applications but are generally obsolete.
Treatment, Control, and Prevention of Foot-and-Mouth Disease
Culling
Animal movement controls
Vaccination
Supportive care (no specific treatment)
In regions that are normally FMD-free, control of the disease is typically attempted by culling all animals on infected premises, and animal movement controls are imposed to decrease the risk of virus spread.
In both normally FMD-free regions and endemic areas, vaccination around outbreaks may be used to limit the spread of the disease.
No treatments for infected animals are available. There is no specific treatment for FMD; however, supportive care may be allowed in countries where FMD is enzootic.
WOAH classifies countries and regions as free from FMD where vaccination is not practiced; free from FMD where vaccination is practiced; suspended status; or no status (8).
The shows areas where prevalence has been high over long periods of time. They are commonly located in economically challenged countries where veterinary services and resources are inadequate to control or eradicate FMD. Europe (in the 1960s and 1970s) and South America (in the 1990s and 2000s) effectively eradicated FMD through coordinated international efforts and large-scale vaccination of cattle with high-quality inactivated vaccines.
However, import controls for animals and animal products have not completely prevented the introduction of FMD into FMD-free areas. These virus incursions into countries or regions where FMD is not enzootic are usually controlled by culling all infected and susceptible animals in infected herds, strict restriction of animal and vehicle movement around infected premises, proper carcass disposal, and environmental disinfection, without the use of vaccines.
FMD-free countries are often reluctant to use vaccination for outbreak control. There is a concern that carrier animals induced by clinically inapparent FMDV infection of vaccinated animals can remain undetected by postvaccination serological testing. Therefore, vaccinated animals, even those without detectable antibodies against nonstructural proteins of FMDV, are often culled as a precautionary measure after the outbreak has been contained. The 2024 revision of the WOAH Terrestrial Animal Health Code, however, allows countries to regain FMD-free status without additional delay even if vaccinated animals remain in the national herd, as long as stringent surveillance requirements are met (8).
Inactivated virus vaccines protect for only 4–6 months against the specific serotypes contained in the vaccine. Billions of doses are used each year and protect animals from clinical illness but not viral persistence in the pharyngeal region (of cattle), and thus vaccinated animals can be carriers of infectious FMD virus. Additionally, distinguishing infected animals from vaccinated animals is difficult unless costly purified vaccines are used. Therefore, vaccination is used primarily in enzootic countries to protect production animals, particularly high-yielding dairy cattle, from clinical illness because slaughter of all at-risk individuals may be economically unfeasible and can cause food shortages.
The production of inactivated vaccines is dangerous and expensive because it requires the cultivation of large quantities of virulent FMDV in sophisticated biocontainment facilities. In experimental settings, many new vaccines without this constraint (eg, viruslike particles, rationally designed immunogenic peptides, synthetic mRNAs, and leaderless or codon-deoptimized viruses, as well as recombinant vector platforms) have delivered promising results; however, none are commercially available.
Rapid disease reporting is essential to control an FMD outbreak in nonendemic countries. During an outbreak, tracing is done through epidemiological inquiries to help find the source of disease introduction. Sequencing of viruses can also identify the source and pathways of spread of closely related viruses.
When mass culling is performed, infected carcasses must be disposed of via incineration, burial, or rendering on or close to the infected premises. Scavengers and rodents should be controlled to prevent mechanical dissemination of virus. Buildings should be cleaned and disinfected with an acidic or alkaline disinfectant, and people who have come into contact with virus must decontaminate their clothing and avoid contact with susceptible animals for a period of time.
In some regions, FMD persistence in wildlife populations, such as African buffalo, can make FMD eradication very difficult. Control measures, such as fencing of wildlife reserves to prevent contact with domestic livestock, have helped limit the spread of virus in certain areas. A twice-yearly vaccination buffer zone in livestock near endemic wildlife reserves could additionally help decrease outbreaks.
A progressive control pathway has been developed by the UN's Food and Agriculture Organization (FAO) and adopted by WOAH to enable countries to improve their FMD control so that the global disease situation will improve (9).
As with all infectious diseases, good on-farm biosecurity is very important for the protection of herds against the introduction of FMDV.
Key Points
Control of FMD can be successfully achieved by strong and well-funded veterinary services, enabling rapid diagnosis and implementation of control measures, including, in some circumstances, the use of vaccines.
In countries that are normally free of FMD, culling of infected animals and those at high risk of getting infected is undertaken; however, public concern about the ethics of mass culling has encouraged the search for improved vaccines that can reliably prevent the carrier state and will allow vaccinate-to-live control policies.
Highly sensitive diagnostic tests for FMD virus and specific antibodies are available; however, the rapid detection of disease incursions into FMD-free areas first and foremost requires awareness by farmers and veterinarians.
For More Information
WOAH Manual of Diagnostic Tests and Vaccines for Terrestrial Animals 2024.
Progressive Control Pathway for Foot-and-Mouth Disease Control. 2nd ed. Food and Agriculture Organization of the UN and European Commission for the Control of Foot-and-Mouth Disease. 2018.
References
Donaldson AI, Gloster J, Harvey LD, Deans DH. Use of prediction models to forecast and analyse airborne spread during the foot-and-mouth disease outbreaks in Brittany, Jersey and the Isle of Wight in 1981. Vet Rec. 1982;110(3):53-57. https://pubmed.ncbi.nlm.nih.gov/7064324/
Brown E, Nelson N, Gubbins S, Colenutt C. Airborne transmission of foot-and-mouth disease virus: a review of past and present perspectives. Viruses. 2022;14(5):1009. doi:10.3390/v14051009
Foot and mouth disease card. World Organisation for Animal Health. Updated January 2025. Accessed February 5, 2026. https://www.woah.org/app/uploads/2021/09/202501-fmd-diseasecard.pdf
Donaldson AI. Risks of spreading foot and mouth disease through milk and dairy products. Rev Sci Tech. 1997;16(1):117-124. doi:10.20506/rst.16.1.1013
Bartley LM, Donnelly CA, Anderson RM. Review of foot-and-mouth disease virus survival in animal excretions and on fomites. Vet Rec. 2002;151(22):667-669. doi:10.1136/vr.151.22.667
Stenfeldt C, Arzt J. The carrier conundrum: a review of recent advances and persistent gaps regarding the carrier state of foot-and-mouth disease virus. Pathogens. 2020;9(3):167. doi:10.3390/pathogens9030167
Alexandersen S, Zhang Z, Donaldson AI, Garland AJ. The pathogenesis and diagnosis of foot-and-mouth disease. J Comp Pathol. 2003;129(1):1-36. doi:10.1016/s0021-9975(03)00041-0
GF-TADS and FMD. Global Framework for the Progressive Control of Transboundary Animal Diseases. Accessed February 5, 2026. https://www.gf-tads.org/fmd/fmd/en/


