



Bilirubin is a degradation product of heme-containing proteins. Most bilirubin (~80%) derives from hemoglobin released from senescent erythrocytes. Hemoglobin is the red pigmented oxygen-binding protein in RBCs. Smaller amounts of bilirubin derive from heme in the bone marrow from disrupted RBC development and heme released from other heme-bearing proteins. The latter sources include:
cytochrome P450 enzymes (metabolic biotransformation enzymes that contain a core-heme molecule, largely found in the liver)
myoglobin (oxygen-binding protein in muscle)
catalases (enzymes catabolizing hydrogen peroxide)
peroxidases (enzymes catalyzing redox-reactions that generate peroxides)
The reticuloendothelial system (RES), an arm of the immune system, mobilizes heme from its parent molecule. This heterogeneous tissue-fixed macrophage population clears particulate and soluble substances from tissues and the systemic circulation. Prominent RES populations reside in the spleen and liver (hepatic Kupffer cells), with smaller numbers in bone marrow and lymph nodes.
Bilirubin derives from the sequential catalytic degradation of heme, mediated by two groups of enzymes: heme oxygenases and biliverdin reductases.
Heme oxygenase resides in the RES (principally the spleen and hepatic Kupffer cells) and initially yields biliverdin, a green pigmented bilirubin metabolite.
Biliverdin is subsequently reduced by biliverdin reductase to an orange-yellow water-insoluble bilirubin pigment: unconjugated bilirubin (UnBr).
UnBr circulates to the liver tightly bound to albumin, and to a lesser extent to high-density lipoproteins. The tight but reversible binding to albumin improves the water solubility of UnBr and limits its extravascular distribution (ie, tissue dissemination and glomerular filtration).
UnBr is cytotoxic; cellular toxicity of UnBr only materializes with concurrent severe hypoalbuminemia and extreme hyperbilirubinemia (~20-fold increase over upper reference range limits) when the molar concentration of bilirubin exceeds its albumin binding partner.
UnBr can provoke neurotoxicity and bilirubin encephalopathy (bilirubin-induced neurologic dysfunction). Kernicterus (macroscopic yellow pigmentation of brainstem tissue) with neurotoxicity is principally encountered in human infants and is exceedingly rare in companion animals. Neonatal kittens injected with bilirubin pigments have been used as an experimental animal model to study kernicterus in humans.

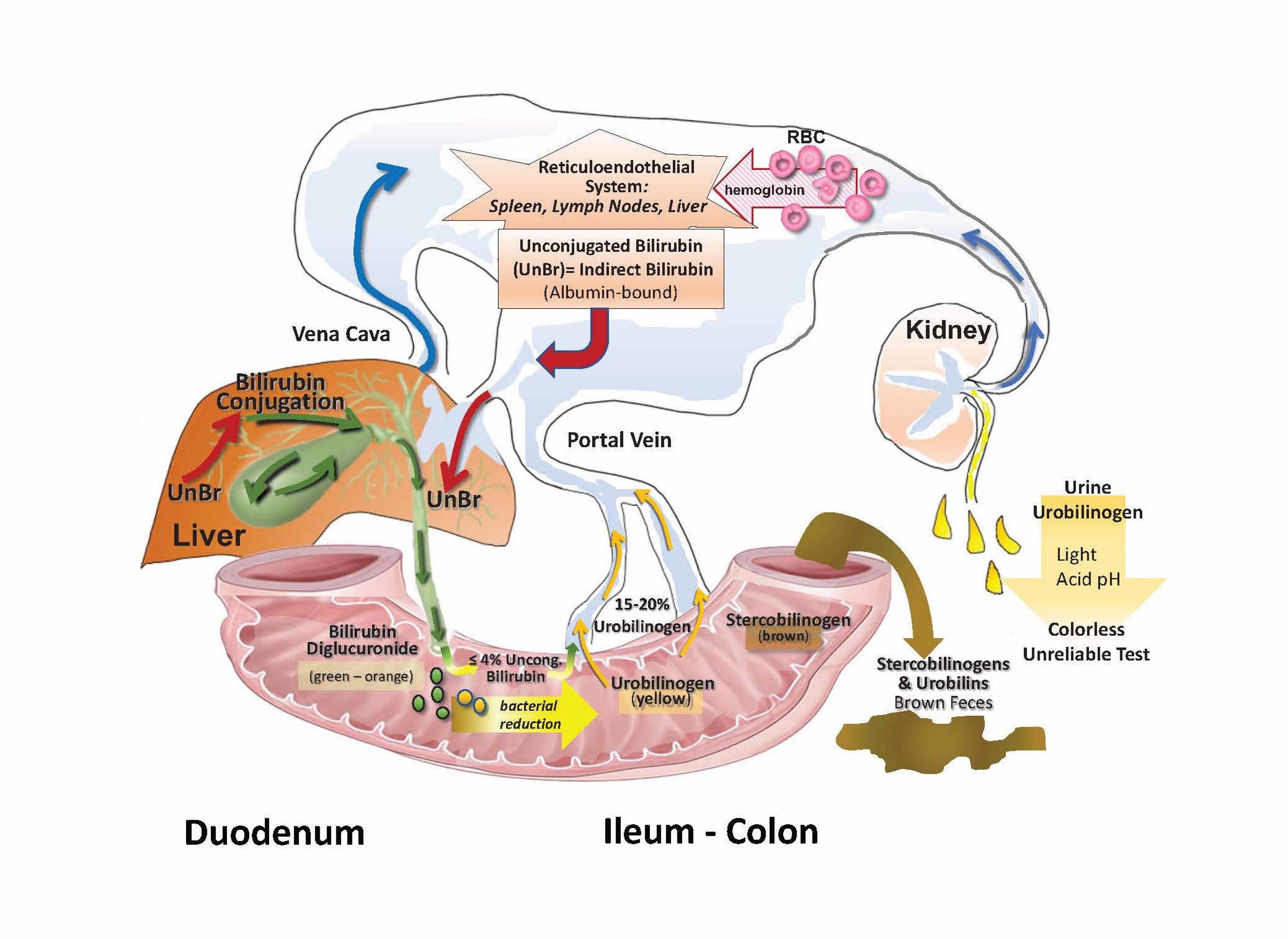
Senescent RBCs are taken up by macrophages in the reticuloendothelial system, where catalytic degradation of hemoglobin leads to release of unconjugated bilirubin (UnBr). As measured by a bilirubin test, the concentration of indirect bilirubin (calculated by subtracting the direct-reacting fraction from the total bilirubin concentration) correlates with the concentration of UnBr. UnBr, bound to albumin, travels in the blood to the liver. Hepatocytes take up UnBr and conjugate it to one or two glucuronic acid residues. Bilirubin travels through the biliary system to the duodenum; most bilirubin in bile is conjugated but some UnBr (up to 4%) in bile or UnBr formed by enteric bacterial deconjugation undergoes enterohepatic recycling. In the terminal ileum and colon, bacterial proteases reduce bilirubin diglucuronide to urobilinogen. Approximately 15% to 20% of urobilinogen is returned to the liver via the portal vein. Urobilinogen remaining in the colon can be converted to stercobilinogen, among other metabolites. Stercobilinogens and urobilins are excreted in feces. Urobilinogen is also excreted in urine.
Courtesy of Dr. Sharon Center.
Bilirubin Plasma Clearance
Plasma clearance of bilirubin reflects the rate of hepatocyte uptake, trivial reflux backward into plasma, and the rate of glucuronide conjugation. At the hepatocyte surface, bilirubin is efficiently extracted as it dissociates from albumin in the space of Disse. (See Anatomy of the Hepatobiliary System.) UnBr is promptly conjugated with glucuronic acid, generating water-soluble bilirubin diglucuronide (or monoglucuronide), denoted as conjugated bilirubin (CBr).
Conjugation facilitates canalicular clearance; there is now a molecular binding partner for this step. Canalicular secretion is considered the rate-limiting factor for bilirubin clearance; CBr contributes ~96%–99% of biliary bilirubin. Bilirubin in bile determines its yellow-green pigmentation.
Upon alimentary discharge with bile, the hydrophilicity and large molecular size of CBr impede its passive resorption. Trivial amounts of UnBr in bile or UnBr formed by enteric bacterial deconjugation undergo some degree of enterohepatic recycling.
In some patients with impaired hepatic bilirubin elimination, oxidation of bilirubin by mixed-function oxidases in liver and elsewhere can assist with bilirubin dissipation (clearance); this is noted by a gradual decline and plateaued hyperbilirubinemia in animals with major bile duct obstruction or pathological ductopenia.
Enteric Bilirubin Metabolites
Once bile enters the alimentary canal, CBr is reduced by colonic bacterial enzymes into a number of urobilinogens. The two major urobilinoids are urobilinogen and stercobilinogen (see ). Initially colorless precursors are transformed into yellow-orange or brown pigmented products upon oxidation to urobilins.
Urobilinogens and their derivatives are partially absorbed from the bowel into the portal circulation and undergo some extent of enterohepatic recycling before elimination in feces or urine.
Animals with hemolytic jaundice can display orange-green to brown fecal pigmentation reflecting extreme formation of urobilinogens and stercobilinogens. In cholestatic jaundice impeding biliary elimination of CBr, renal clearance may account for 50%–90% of bilirubin elimination (orange urine pigmentation, despite lack of urine concentration). With unconjugated hyperbilirubinemia, biliary clearance is the only route of UnBr elimination.
Urinary Urobilinogen
Urine urobilinogen can be decreased, normal, or increased in patients with liver disease. Increased urinary urobilinogen may reflect increased bilirubin production, decreased bilirubin clearance (eg, in advanced liver disease or portosystemic shunting (acquired or congenital in animals faced with hyperbilirubinemia), or escalated enteric bacterial generation of urobilinogen due to constipation or enteric bacterial overgrowth. Absence of urinary urobilinogen accompanied by acholic feces (no brown pigmentation) in a jaundiced animal implies complete bile duct obstruction or pathological ductopenia as underlying causes.
Yet this relationship is not inviolable because enteric bleeding and compensatory expression of bilirubin transporters on the intestinal wall can flux enough bilirubin into the alimentary canal to generate fecal pigmentation (urobilins). The discordant absence of fecal urobilins or urinary urobilins in the presence of marked hepatobiliary jaundice (not caused by bile duct obstruction or extreme ductopenia) can also occur. Examples include enteric dysbiosis caused by indiscriminate chronic oral antibiotic administration (suppressing enteric generation of pigmented urobilins), increased renal tubular urobilin resorption, or urobilinogen instability in acid urine stored with light exposure.
Additionally, standard clinical assessments of urine urobilinogen cannot distinguish between normal and subnormal concentrations. Consequently, urinary urobilinogen assessment has limited diagnostic value.
Persistent Jaundice
Because bilirubin can form strong bonds with elastin in skin and sclera, jaundice may persist despite normalized plasma bilirubin concentrations. This is most often encountered in animals with prolonged conjugated hyperbilirubinemia (eg, weeks of major bile duct obstruction or severe jaundice caused by hemolytic anemia) where delta-bilirubin may accumulate.
Delta-bilirubin is CBr that has formed an irreversible covalent bond with albumin. Jaundice associated with delta-bilirubin lingers because it cannot be cleared by liver or kidney (due to the persistent albumin bond) with slow clearance reflecting the ~10–14 day half-life of albumin. In the absence of delta-bilirubin, healthy dogs have a CBr clearance of ~2–4 hours.
Jaundice associated with delta-bilirubin may lack detectible urine bilirubin on dipstick surveillance. Although it is not important to measure delta-bilirubin, its presence may confound appraisal of treatment response.
Another phenomenon associated with delta-bilirubin is that some patients exhibit an increase in plasma bilirubin concentration after plasma or albumin transfusion, reflecting bilirubin flux from tissues to the intravascular compartment. Accrual of delta-bilirubin additionally contributes to the poor clinical utility of van den Bergh bilirubin fractionation in that delta-bilirubin yields a direct diazo reaction.
Irrespective of delta-bilirubin accrual, all jaundiced animals accumulate CBr in tissues and fluids that house high albumin concentrations. Measurement of bilirubin concentration in abdominal effusions is often explored to exonerate consideration of bile peritonitis as a cause of jaundice. In the absence of bile peritonitis, effusion bilirubin concentrations are coordinate with plasma bilirubin concentrations, contrasting with extreme bilirubin concentrates in effusions associated with bile peritonitis.





